El seleni (Se) és un micronutrient essencial que s’aconsegueix mitjançant la dieta i que té propietats antioxidants i antiiflamatòries, regulades a través de les selenoproteïnes. Per tant, el Se és un element essencial pel bon funcionament de les selenoproteïnes. [11][12]
La dosi òptima de seleni és de 55μg adult/dia. Tot i que és rar trobar casos de toxicitat o deficiència extrema d’aquest oligoelement, s’ha demostrat que canvis en els nivells de seleni poden afectar significativament la salut humana [13] [14]. Els efectes atribuïts a canvis dels nivells de Se són multifactorials i inclouen la salut cardiovascular, el metabolisme hormonal de la tiroide, la inflamació i la funció immune, així com la protecció contra el dany oxidatiu i la neurodegeneració, el càncer i la infecció viral. [15] Sabies què? Després d’un elevat percentatge de morts de nens adolescents i dones joves a Keshan (Xina), l’any 1979 es va descobrir que tots els habitants d’aquella regió, uns 50 milions, tenien una carència de seleni en sang, cabell i orina. Després de la suplementació amb 0,3mg de seleni per setmana la malaltia (Kashin-Beck) es va poder eradicar quasi per complet.
El seleni en Ictalurus punctatus
Deficiències de seleni i vitamina E en Ictalurus punctatus s’han relacionat amb supressió del creixement, anèmia, miopatia severa, diàtesis exsudativa i mort. La deficiència d’un dels dos elements, en canvi, no produeix cap deficiència. Per tant, es pot establir que hi ha una relació significant entre el Se i la vitamina E en la dieta d’Ictalurus punctatus.
2. Les selenoproteïnes
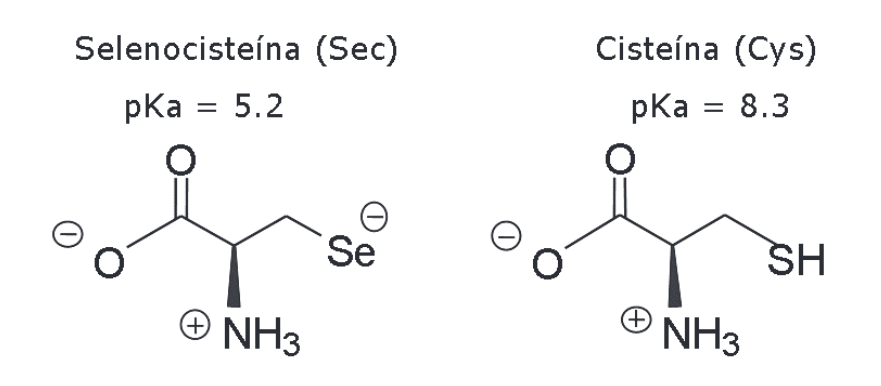
Les selenoproteïnes són una família de proteïnes que contenen en la seva estructura primària una selenocisteïna (Sec) , l'aminoàcid 21a en el codi genètic. Sec és un anàleg de la cisteïna en la qual la cadena lateral que conté sofre (SH) se substitueix per una cadena lateral que conté seleni (Se-) [16].
Figura 1. Fórmula de la selenocisteïna (Sec) i de la cisteïna (Cys). Consulta l'article complet aquí.
Extensament classificades com a antioxidants, les selenoproteïnes actuen sobre una gran varietat de substrats i tenen múltiples funcions [14]. Entre elles controlen l’estat redox cel·lular, protegeixen contra el dany oxidatiu i l’estrès, el metabolisme tiroïdal, la mobilització del Ca2+ intracel·lular [17][18], el plegament de les proteïnes [19], el transport de seleni [20], i fins i tot actuen com a intermediàries en la síntesi de Sec. [21][22]
Sec està codificada pel codó UGA, un dels tres codons STOP per la traducció de mRNA en la resta de gens. El reconeixement del codó UGA com a lloc d’inserció de Sec enlloc d’una senyal de parada requereix la presència d’un element SECIS (SelenoCysteine insertion sequence) en la regió 3’ UTR del mRNA de la selenoproteïna i un tRNA-selenocisteil únic. A diferència dels 20 aminoàcids estàndars, la Sec es biosintetitzada a partir de la serina en el seu tRNA.
Fins ara, s’ha descrit un gran número i varietat de selenoproteïnes. En concret, en vertebrats s’han definit 45 famílies, mentre que en el genoma humà 25. La majoria de selenoproteïnes van ser descobertes per aproximacions bioinformàtiques buscant elements SECIS.
3. Biosíntesi de les selenoproteïnes
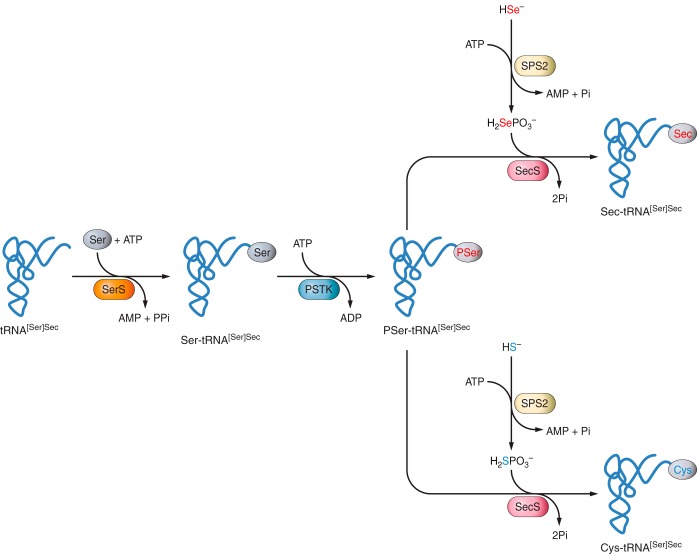
El seleni obtingut de la dieta ha de ser convertit a selenur (Se2-) per tal d’incorporar-se a les selenoproteïnes. Les formes inorgàniques com el selenit (SeO32-) són convertides a Se2- gràcies a les glutaredoxines i les tioredoxines, mentre que les formes orgàniques són metabolitzades per la seleocisteïna liasa (Scly) o per trans-selenació per obtenir també Se2- . Posteriorment, la selenofosfat sintetasa 2 (SPS2) dóna lloc al selenofosfat (SePo33-) i la selenocisteïna (Sec) és incorporada a través del seu tRNA únic (tRNA [Ser]Sec). tRNA [Ser]Sec decodifica el codó UGA com a Sec en lloc d’un codó STOP i així preveu la terminació prematura, però perquè això passi cal que SECIS recodifiqui el codó. [23][24]
Figura 2. Mecanisme de biosíntesi de Sec en eucariotes. En la via de síntesi de Sec, la fosfoseril-tRNA quinasa (PSTK) serveix com a intermediària per fosforilar la Ser-tRNA [Ser]-sec. La selenofosfatasa (H2SePO3-) generada per la SPS2 i l’ATP es fa servir com a donadora de Se per la SecS. En la part inferior dreta es veu la síntesi de novo de Cys. Imatge extreta de : Labunskyy V, Hatfield D, Gladyshev V. Selenoproteins: Molecular Pathways and Physiological Roles. Physiological Reviews. 2014;94(3):739-777. Article disponible aquí.
SECIS és una estructura tridimensional d’ARN en forma de loop que permet el reclutament de la maquinària de traducció i evita que es codifiqui pel codó STOP. Es troba en l’extrem 3’-UTR del gen de la selenoproteïna en eucariotes i downstream del codó UGA en procariotes.
El procés d’incorporació de la selenocisteïna comença quan l’element SECIS recluta la proteïna SECIS Binding Protein (SBP2). Això permet la unió del factor d’elongació específic de selenoproteïnes (eEFSec), que alhora atrau el selenocisteinil-tRNA (tRNA [Ser]Sec) mediant la inserció de Sec en resposta a un codó UGA. A partir d’aquí la traducció continua fins que el ribosoma reconeix un nou codó de terminació. S’han identificat altres proteïnes com la proteïna ribosomal L30 (RPL30) que forma part de la maquinària d’inserció de Sec.
Figura 3. Mecanisme d’inserció de Sec en eucariotes. La figura mostra els factors coneguts necessaris per la incorporació de Sec a les proteïnes en resposta al codó UGA. A més, es mostren els elements que poden influenciar l’eficiència de la inserció com la proteïna ribosomal L30, el factor eIF4a3 i nucleolina (veure el text per més detalls). Imatge extreta de : Labunskyy V, Hatfield D, Gladyshev V. Selenoproteins: Molecular Pathways and Physiological Roles. Physiological Reviews. 2014;94(3):739-777. Disponible aquí.
4. Evolució de les selenoproteïnes en vertebrats
Les tècniques de next-generation sequencing han permès en els darrers anys caracteritzar el genoma de moltes espècies ràpidament. Aquestes eines també han estat útils per l'anàlisi del selenoproteoma, però tot i això encara hi ha moltes espècies per les quals no s’ha estudiat les selenoproteïnes. Cal remarcar que l’estudi de les selenoproteïnes és interessant per conèixer els processos evolutius.
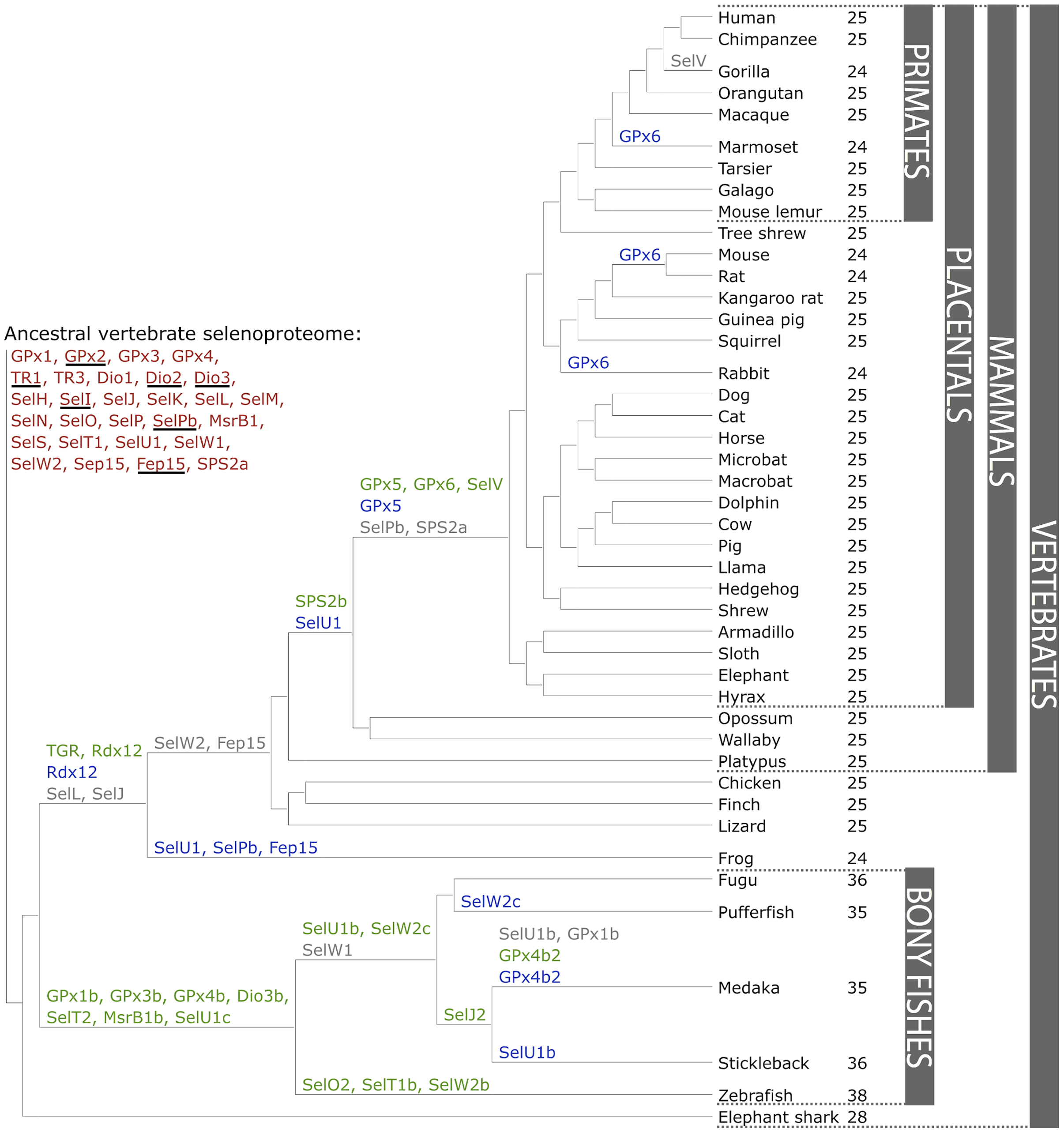
Com es pot veure en la imatge, al llarg de l’evolució el selenoproteoma de les espècies també ha anat variant. En l’arbre filogenètic podem veure els punts on s’han produït duplicacions (indicat en verd), on s’han produït pèrdues d'un selenoproteïna en particular (en gris), o substitucion de Sec per Cys (en blau).
Si ens fixem en l'arbre filogenètic de les diferents famílies de selenoproteïnes en els vertebrats, podem veure que l'espècie que estem estudiant, Ictalurus punctatus , no figura a la classificació dels peixos ossis, de manera que per predir les diferents selenoproteïnes en aquest genoma basarem la nostra recerca en una altra espècie que es troba evolutivament molt propera com és el peix zebra (Danio rerio).
Pel que fa a l’evolució del selenoproteoma en vertebrats, sabem que els organismes aquàtics tenen selenoproteomes més grans en comparació als organismes terrestres. D’aquesta manera, l’espècie que conté el màxim nombre de selenoproteïnes és el peix zebra. A més, s’ha descrit que els gens que codifiquen per algunes selenoproteïnes es dupliquen en els peixos ossis (per exemple GPx1b, GPx3b i GPx4b entre d’altres). En canvi, algunes selenoproteïnes com SelL i SelJ es van perdre quan es va donar la colonització del medi terrestre.
Tot això comporta que en l’estudi d’Ictalurus punctatus esperem trobar un gran número de selenoproteïnes (similar al de Danio rerio) i duplicacions que dificultaran la interpretació dels resultats de l’anàlisi. Serà extens i complicat, és per això que intentarem automatitzar el procés per facilitar la feina. [19]
Figura 4. Evolució del selenoproteoma dels vertebats. En vermell s’indica el selenoproteoma ancestral; les proteïnes subrallades corresponen a les úniques que s’han trobat en vertebrats. En verd es mostren les duplicacions de selenoproteïnes; en gris les pèrdues i en blau les substitucions de selenocisteïna (Sec) a cisteïna (Cys). El número situat al costat de cada espècie correspon al nombre de selenoproteïnes trobades. Com es pot veure el peix zebra és l’espècie amb més selenoproteïnes identificades, amb un total de 38.
Imatge extrateta de: Mariotti M, Ridge PG, Zhang Y, Lobanov A, Pringle TH, Guigó R, Hatfield DL, Gladyshev VN. Composition and evolution of the vertebrate and mammalian selenoproteomes. PLoS One [revista a Internet] 2012; 7(3): e33066. Disponible aquí.
5. Selenoproteïnes conegudes
GPx
La família glutatió peroxidases (GPx) està composta de múltiples enzims (8) que estan involucrats en la senyalització del peròxid d’hidrogen (H2O2), la detoxificació de les hidroperoxidases i en el manteniment de l’homeostasi redox cel·lular. GPx1: és la primera que es va identificar. S’expressa en totes les cel·lules, però sobretot en les del fetge i del ronyó. Reacciona amb el peròxid d’hidrogen i hidroperoxidases modulant-ne moltes de les funcions. També té un paper protector enfront l’estrés oxidatiu.
GPx2: Forma part de l’epitel·li gastrointestinal i actua com a barrera per evitar la toxicitat dels hidroperòxids ingerits a través de la dieta.
GPx3: Principalment es troba en el ronyó i en el plasma. No es coneix la seva funció però s’utilitza com a marcador per conèixer l’estat del seleni.
GPx4: Es troba en molts tipus cel·lulars i teixits. Catalitza la reducció de peròxids lipídics.
GPx5: Es troba a l’esperma. Es troba present en mamífers però no en els peixos ossis.
GPx6: Només es troba a l’epitel·li olfactori durant el desenvolupament embrionari. Igual que GPx5 no està present en els peixos ossis.
GPx7: és una proteïna secretada.
GPx8: es troba localitzada a la membrana cel·lular.
DI
La família iodotironina deiodinases (DI) està formada per tres proteïnes paràlogues (DI1, DI2 i DI3), involucrades en la regulació de l’activitat de l’hormona tiroidea. Presenten diferent localització subcel·lular: DI1 i DI3 estan localitzades en la membrana plasmàtica mentre DI2 es troba en el reticle endoplasmàtic.
L’hormona tiroidea produïda per la tiroide és secretada majoritàriament en la seva forma inactiva o T4. Aquesta protohormona pot ser convertida en hormona activa o T3 gràcies a la desiodació de l’anell extern que duen a terme a través d’una reacció DI1 i DI2. DI3, en canvi, s’encarrega d’inactivar l’hormona. Així doncs, les desiodinases tenen un paper important mantenint els nivells i l’activitat tiroidea.
TR o TXNRD
La família tioredoxina reductases (TR) són oxidoreductases que juntament a la tioredoxina (Trx) estan involucrades en la regulació redox dels processos cel·lulars. En mamífers existeixen tres isoformes de la TR: TR1: proteïna amb 6 isoformes diferents que es formen per splicing alternatiu i extensió de l’extrem NH2 terminal. La seva funció és reduir NADPH depenent de la Trx1 i participa en processos com la regulació de factors de transcripció, apoptosis, etc. Es troba al citosol i al nucli. No es troba en peixos ossis. TR2: Forma mitocondrial important en la compactació de l’oxigen reactiu. TR3: proteïna amb múltiples isoformes implicada en la reducció de la tioredoxina mitocondrial (Trx2), motiu pel qual es troba al mitocondri. Està implicada en la defensa contra l’estrés oxidatiu.
Sel W, T, H i V
Són selenoproteïnes que pertanyen a la família Rdz. Aquesta família es caracteritza per tenir un motiu Cys-x-x-Sec conservat en la seva seqüència. Sel W: és una de les selenoproteïnes més abundants en mamífers. Es localitza al citosol i s’expressa en nivells elevats al múscul i cervell. Sel T: es localitza predominantment en el RE i l’aparell de Golgi, i s’expressa ubiquitinada durant el desenvolupament embrionari i en teixits adults. Té un paper fonamental en la regulació de l’homeostasis del Ca2+ i en la funció neuroendocrina. Participa en l’adhesió cel·lular i en l expressió de gens oxidoreductases.
Sel H: presenta un patró de localització subcel·lular únic, en el nuclèol. L’expressió d’aquesta proteïna és molt baixa en teixits adults però en canvi és elevada durant el desenvolupament embrionari. Té la capacitat d’unir-se a seqüències de DNA que contenen heat shocks i elements de resposta a l’estrès. Presenta activitat glutatió peroxidasa i està implicada en la regulació de la transcripció de gens involucrats en la síntesis de novo de glutatió.
Sel V: es localitza en la placenta d’algunes espècies de mamífers. Sel V és un homòleg proper de la Sel W però presenta una mida major a causa d’un domini NH2 terminal addicional. La seva funció encara no es coneix, però creuen que podria estar implicada en la reproducció masculina.
Sel M i Seleno E (Fep15)
La selenoproteïna M (Sel M) és una selenoproteïna que es troba en tots els vertebrats, que s’expressa majoritàriament al cervell i que comparteix un 31% d’identitat de la seqüència amb Sel15 (homòlegs distants). Fep15 O SELENO E es va generar durant la duplicació del genoma i va evolucionar en concret per duplicació de Sel M.
Sel 15
La selenoproteïna 15 (Sel15) és una de les selenoproteines ancestrals i es troba en tots els vertebrats. S’ha proposat que protegeix de l’efecte cancerigen del Se de la dieta i que en el reticle endoplasmàtic regula l’homeostasi i el plegament de les proteïnes. Presenta uns nivells d’expressió màxims a la pròstata, fetge, ronyó i testicles.
Sel R (MSR)
Els enzims Mrs (metionina sulfoxida reductasa) són enzims que tenen com a funció reparar els residus oxidats de metionina (Met) ja que la Met és un aminoàcid susceptible a les espècies reactives d’oxigen de l’organisme. Per tant, catalitzen la conversió de metionina sulfoxida a metionina.
Sel P
La selenoproteïna P (Sel P) és una proteina ancestral que es troba en tots els vertebrats. És una proteïna que se secreta i que conté el 50% del seleni en plasma, i s’encarrega de mantenir les reserves de Se als teixits perifèrics. És la única selenoproteïna amb múltiples residus Sec i presenta 4 isoformes. Es creu que podria tenir propietats antioxidants.
Sel O
La selenoproteïna O (Sel O) és una selenoproteïna mitocondrialredox-activa que es troba en tots els vertebrats i que s’expressa en diferents teixits.
Sel I
La selenoproteïna I (Sel I) és una selenoproteïna ancestral que es troba en tots els vertebrats. És una de les últimes selenoproteïnes descobertes. És troba en el RE com a proteïna transmembrana i es creu que podria participar en la biosíntesi dels fosfolípids.
Sel S i Sel K
Les selenoproteïnes S i K (SelS i SelK) es poden agrupar dins la mateixa família ja que ambdues tenen un domini transmembrana, presència d’un segment ric en glicina, i aminoàcids carregats positivament. Tot i això no tenen similituds en la seqüencia.
SelS: selenoproteïna ancestral present en tots els vertebrats. Es troba al RE i a la membrana cel·lular. Té una funció important en resposta a l’estrès causat per les proteïnes mal plegades (unfolded protein response o UPR) formant part del sistema de degradació proteica associada al ER (ERAD). Proporciona protecció contra ROS. SelK: com la Sel S és ancestral i es troba present en tots els vertebrats. Es troba al RE i a la membrana plasmàtica. Està involucrada en la degradació de proteïnes glicosilades associades al sistema ERAD. També participa en la facilitació del flux de calci a les cèl·lules immunitàries.
Sel N
La selenoproteïna N (Sel N) és ancestral i es troba en tots els vertebrats. Es localitza al RE i s’expressa majoritàriament en el desenvolupament embrionari i en el creixement dels músculs, cor, pulmons i placenta. Una mutació en el seu gen (SEPN1) causa desordres musculars coneguts amb el nom de miopaties de tipus SEPN1.
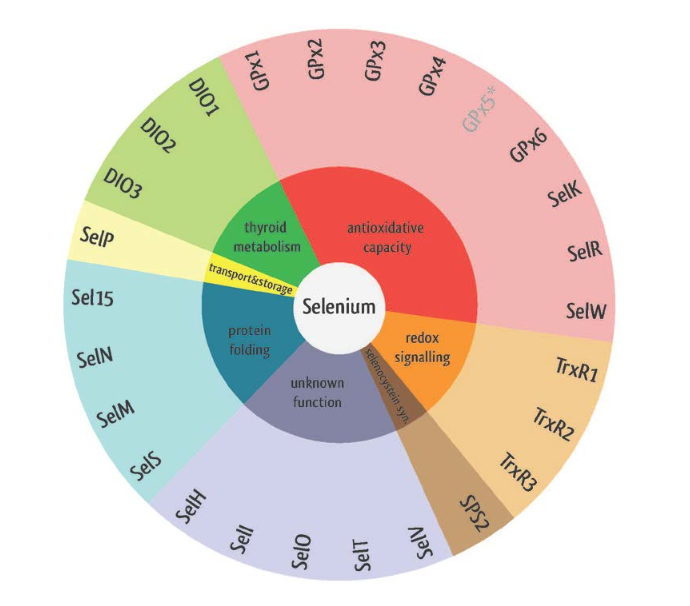
Figura 5. Classes de selenoproteïnes i les seves funcions resumides. Imatge extreta de: Benstoem C, Goetzenich A, Kraemer S, Borosch S, Manzanares W, Hardy G et al. Selenium and Its Supplementation in Cardiovascular Disease—What do We Know?. Nutrients. 2015;7(5):3094-3118. Disponible aquí.
6. Selenoproteïnes conegudes de maquinària
SPHS
La selenofosfatasa sintetasa 2 (SEPHS, també anomenada SPS) és la única entre els components de la maquinària de biosíntesi de Sec que a vegades és una selenoproteïna per ella mateixa. SPS catalitza la síntesi del selenofosfat a partir d’ATP i selenit. El selenofosfat és el donador de seleni per la síntesi de Sec. Les proteïnes SPS estan conservades des dels bacteris fins els humans amb un 30% d’identitat. Existeixen dos gens SPS paràlegs:
- SEPHS1: És una selenoproteïina
- SEPHS2: No es una selenoproteïna.
eEFsec
El factor d'elongació eucariò,tic o eEFsec en eucariotes (i Sel B en procariotes) promou la incorporació de selenocisteïna en les selenoproteines amb un mecanisme que encara està poc entès.
PTSK
La fosfoseril-tRNA quinasa (PTSK) és una proteïna que està conservada en eucariotes i arqueobacteris, fet que suggereix un paper important en la síntesi o regulació de les selenoproteïnes. Catalitza la reacció de seril-tRNA [Ser]Sec a la forma fosforilada fent servir una molècula d’ATP.
SBP2
SBP2 prmou la incorporació de Sec associant-se amb l’element SSECIS i reclutant el complex eEFsec-selenocisteil-tRNA[Ser]Sec.
SECp43
SECp43 es requereix per a la maduració de Sec-tRNA.
SecS
SecS uneix la molècula de selenofosfat a la serinda portada pel seril-RNA
SBP
La Secis Binding protein reconeix l’element SECIS i recluta eEFSec i altres factors per recodificar el codó UGA