Introducció
Aquest apartat tracta dels protists i, més concretament, dels organismes protists analitzats al llarg de tot aquest projecte de recerca. També podreu trobar un arbre filogenètic dels protistes que nosaltres hem analizat i altres estudiats en anys anteriors.
A més a més, contés una introducció general a les selenoproteïnes, així com també l'explicació de les diferents famílies de selenoproteïnes que nosaltres hem analitzat (EhSEP2,Sel O, Sel S).
Protists
Els protists són un grup molt divers d’organismes eucariotes, la majoria dels quals són microscòpics. Aquest grup comprèn, entre d’altres, algues, eucariotes i protozous.
Els protists són membres dels Eukarya, el tercer domini en l’arbre de la vida. Les cèl·lules eucariotes són característiques tant dels protists com dels organismes pluricel·lulars complexes presents en els altres 3 regnes eucariotes.
Els protists són descendents dels primers eucariotes, mentre que els animals, els fongs i les plantes van sorgir d’ancestres protistes. Tot i això, la majoria són de vida lliure però alguns poden formar associacions simbiòtiques amb diversos organismes no relacionats. Aquestes associacions són el mutualisme, el comensalisme i el parasitisme.Els protists varien en el pla corporal, en els mètodes de moviment, en la seva nutrició i en el mètode de reproducció.
Mida
La mida varia de manera considerable dins del regne protista, des dels protozous microscòpics fins a algunes algues gegants. La majoria d'organismes protists són unicel·lulars, és a dir, cada cèl·lula dóna lloc a un organisme complex capaç de realitzar totes les funcions necessàries per a la vida. Ara bé, també trobem protists formant colònies (agrupacions laxes de cèl·lules), altres són cenocítics (masses multinucleades de citoplasma) i altres són pluricel·lulars (composats per moltes cèl·lules). D’altra banda, i a diferència dels animals, els protists pluricel·lulars tenen formes corporals relativament simples i sense teixits especialitzats.
Finalment, remarcar que la majoria dels protistes són mòbils en algun moment del seu cicle vital i poden presentar diversos mètodes de locomoció.
Nutrició
Els mètodes per obtenir nutrients també varien molt. Per exemple, els flagel·lats s’alimenten per filtració mentre que altres protists poden fagocitar bactèries. Les algues, per exemple, són autòtrofes i fan la fotosíntesis mentre que altres protists obtenen l’aliment per absorció.
Ambient
Gairebé tots els protists són aquàtics i viuen a l’oceà i en estanys d’aigua dolça, llacs i rierols. Els protists terrestres estan restringits a llocs humits, com en el sòl, en esquerdes o enmig de la fullaraca.
Reproducció
Dins del regne protista la reproducció també és molt variada. Gairebé tots es reprodueixen asexualment però n’hi ha d’altres que es reprodueixen sexualment. Tot i això, la majoria de protists no desenvolupa òrgans reproductors pluricel·lulars ni forma embrions.
Protists estudiats
Acanthopagrus taiwanensis
Acanthopagrus taiwanensis és un peix teleosti de la família dels espàrids i de l'ordre dels perciformes. Aquest es troba restringit taxonòmicament a les costes de Taiwan i zones de clima temperat.| Regne | Protist |
| Fílum | Chordata |
| Classe | Actinopterygii |
| Ordre | Perciformes |
| Família | Sparidae |
| Gènere | Acanthopagrus |
| Espècie | A. taiwanensis |
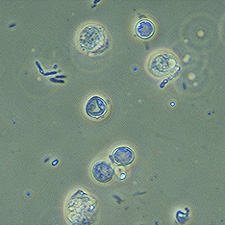
Blastocystis hominis
Blastocystis hominis és un protozou que infecta a l’espècie humana causant diarrea. Aquest es transmet de forma fecal-oral i, en el cas de la transmissió animal-humà, el reservori més conegut de B.hominis són els porcs i els gossos.| Regne | Protist |
| Fílum | Heterokontophyta |
| Classe | Blastocystea |
| Ordre | Blastocystida |
| Família | Blastocystidae |
| Gènere | Blastocystis |
| Espècie | B.hominis |
Per a més informació podeu veure l'entrada de Viquipèdia per a Blastocystis hominis que hem creat l'11 març 2011 a les 13:40.
Capsaspora owczarzaki
Capsaspora owczarzaki és un eucariota uncel·lular que estableix relacions de simbiosi amb el cargol. Algunes característiques determinants d’aquesta espècie són la capacitat de desenvolupar-se a l’interior de l’hemolimfa del cargol, ingerir larves tremàtodes i reproduïr-se per fissió asexual. A més a més, C.owczarzaki és l’hoste intermediari de l’espècie Schistosoma, un platihelmint que és l’agent causant de l’esquistosomiasis.| Regne | Protist |
| Fílum | Choanozoa |
| Classe | Ichthyosporea |
| Grup | Fungi/Metazoa |
| Família | Fungi/Metazoa incertae sedis |
| Gènere | Capsaspora |
| Espècie | C.owczarzaki |
Per a més informació podeu veure l'entrada de Viquipèdia per a Capsaspora owczarzaki que hem creat l'11 març 2011 a les 15:30.
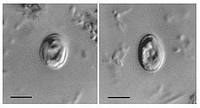
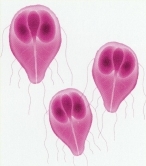
Cryptosporidium muris
Cryptosporidium muris és un protozou que viu com a paràsit. Tradicionalent aquest no ha sigut considerat un patogen humà, però recentment s’han diagnosticat casos humans infectats per C.muris, per això es creu que podria tractar-se d’un patògen zoonòtic emergent capaç d’infectar a l’espècie humana.| Regne | Chromalveolata |
| Fílum | Apicomplexa |
| Classe | Conoidasida |
| Ordre | Eucoccidiorida |
| Família | Cryptosporidiidae |
| Gènere | Cryptosporidium |
| Espècie | C.muris |
Per a més informació podeu veure l'entrada de Viquipèdia per a Cryptosporidium muris que hem creat l'11 març 2011 a les 15:35.
Cryptosporidium parvum
Cryptosporidium parvum és el protozou causant de la criptosporidosis, una malaltia que afecta al tracte intestinal d’humans. C.parvum és considerat un dels patogen aquàtics més importants present en els països desenvolupats, ja que és capaç de sobreviure davant de pràcticament tots els nivells de cloració de les aigües.| Regne | Chromalveolata |
| Fílum | Apicomplexa |
| Classe | Conoidasida |
| Ordre | Eucoccidiorida |
| Família | Cryptosporidiidae |
| Gènere | Cryptosporidium |
| Espècie | C.parvum |
Per a més informació podeu veure l'entrada de Viquipèdia per a Cryptosporidium parvum que hem creat l'11 març 2011 a les 15:51.
Cyanidioschyzon merolae
Cyanidioschyzon merolae és una alga vermella unicel·lular que viu en ambients molt àcids. Aquesta alga té un únic cloroplast i una única mitocondria, però careix de vacuol i paret cel·lular.| Regne | Protist |
| Fílum | Rhodophyta |
| Classe | Cyanidiophyceae |
| Ordre | Cyanidiales |
| Família | Cyanidiaceae |
| Gènere | Cyanidioschyzon |
| Espècie | C.merolae |
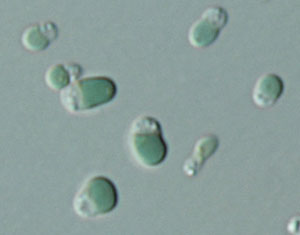
Per a més informació podeu veure l'entrada de Viquipèdia per a Cyanidioschyzon merolae que hem creat l'11 març 2011 a les 15:54.
Ectocarpus siliculosus
Ectocarpus siliculosus és una alga marró filamentosa pròpia de zones temperades i costaneres. Aquesta comparteix algunes característiques amb les plantes terrestres, així com la capacitat fotoautotròfa i la presència de cel·lulosa en la paret cel·lular.El genoma d’E.siliculosus és l’únic genoma d’algues multicel·lulars completament caracteritzat i anotat i, com a tal, E.siliculosus és el model d’elecció alhora d’estudiar aquest tipus d’algues.
| Regne | Chromalveolata |
| Fílum | Heterokontophyta |
| Classe | Phaeophyceae |
| Ordre | Ectocarpales |
| Família | Ectocarpaceae |
| Gènere | Ectocarpus |
| Espècie | E.siliculosus |
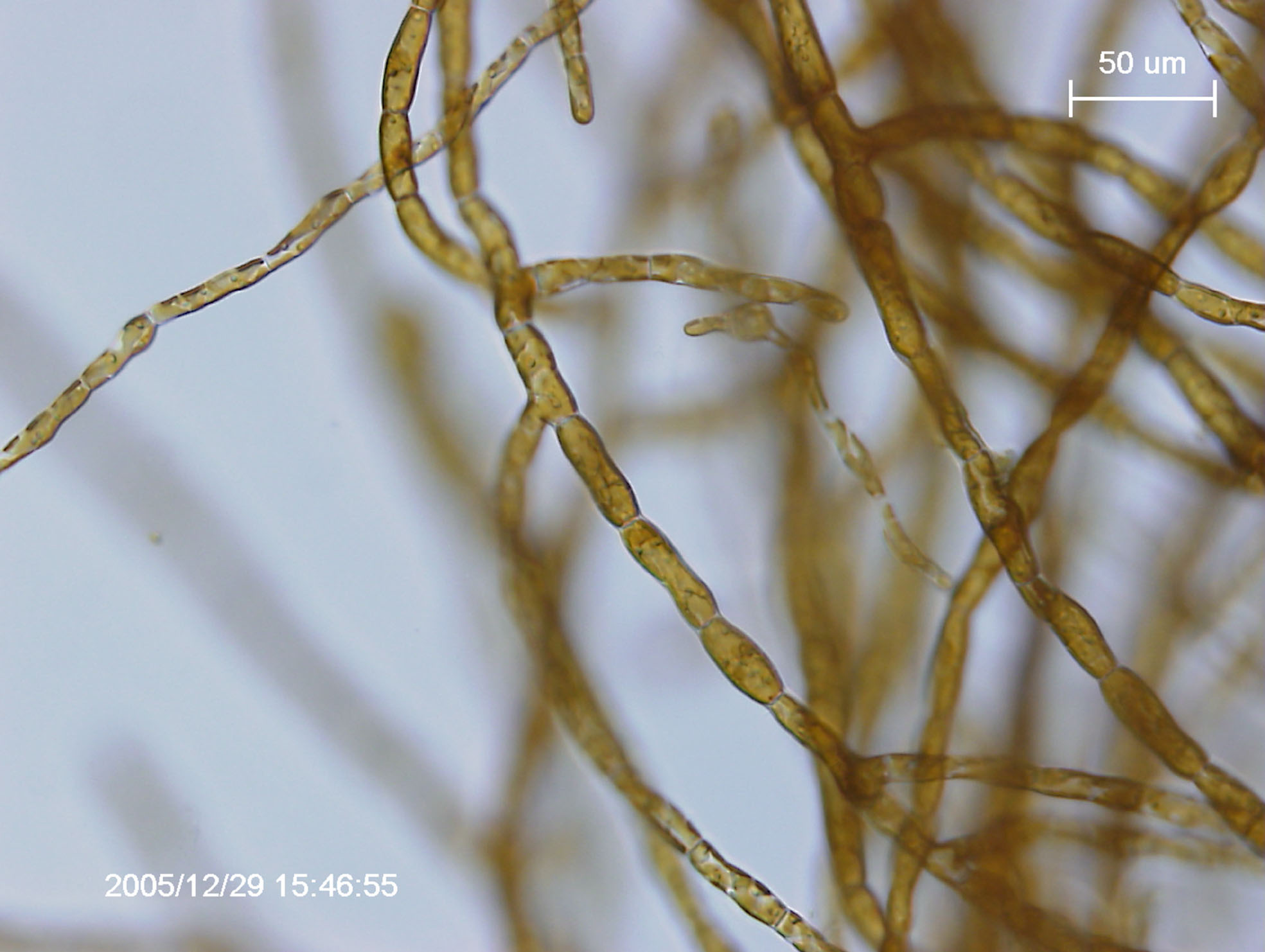
Per a més informació podeu veure l'entrada de Viquipèdia per a Ectocarpus siliculosus que hem creat l'11 març 2011 a les 15:57.
Hyaloperonospora arabidopsidis
Hyaloperonospora arabidopsidis és un paràsit obligat que infecta al model de plantes arabidopsis. Aquesta espècie és altament resistent a la gran diversitat d’hostes, així com també a gran part de proteïnes patògenes avirulents.La seqüènciació del genoma d’aquest organisme ha permés fer anàlisis bioinformàtics amb l’objectiu d’esbrinar els mecanismes defensius i manipular altres processos cel·lulars de la cèl·lula hoste.
| Regne | Chromalveolata |
| Fílum | Heterokontophyta |
| Classe | Oomycetes |
| Ordre | Peronosporales |
| Família | Peronosporaceae |
| Gènere | Hyaloperonospora |
| Espècie | H.arabidopsidis |
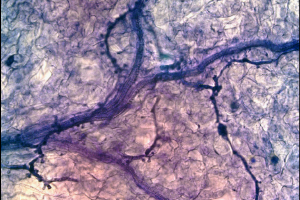
Per a més informació podeu veure l'entrada de Viquipèdia per a Hyaloperonospora arabidopsidis que hem creat l'11 març 2011 a les 16:00.
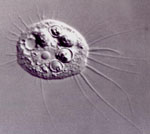
Naegleria gruberi
Naegleria gruberi és una ameba flagelada no patògena que destaca per la seva capacitat de canviar de la forma ameboide a la flagelada. En condicions normals, N.gruberi creix i es divideix com una amoeba, però en condicions de falta de nutrients N.gruberi esdevé un flagelat, tot desenvolupant un esquelet de microtúbuls. No obstant, aquest estadi flagelat és transitori.El genoma de N.gruberi és clau a l'hora d’estudiar processos evolutius d’ameba.
| Regne | Protist |
| Fílum | Percolozoa |
| Classe | Heterolobosea |
| Ordre | Schizopyrenida |
| Família | Vahlkampfiidae |
| Gènere | Naegleria |
| Espècie | N.gruberi |
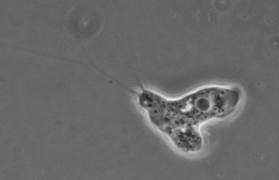
Per a més informació podeu veure l'entrada de Viquipèdia per a Naegleria gruberi que hem creat l'11 març 2011 a les 16:03.
Phaeodactylum tricornutum
Phaeodactylum tricornutum és una alga diatomea i és l’única espècie del gènere Phaeodactylum. Aquesta alga adopta diferents tipus morfològics, fet que la fa útil per estudiar alteracions en la forma cel·lular, els quals poden ser estimulats per canvis en les condicions ambientals. A més a més, P.tricornutum pot créixer en absència de silicona, fet que proporciona noves eines en el camp de la fabricació de nanoinstruments de silicona.| Regne | Chromalveolata |
| Fílum | Heterokontophyta |
| Classe | Bacillariophyceae |
| Ordre | Naviculales |
| Família | Phaeodactylaceae |
| Gènere | Phaeodactylum |
| Espècie | P.tricornutum |
Per a més informació podeu veure l'entrada de Viquipèdia per a Phaeodactylum tricornutum que hem editat l'11 març 2011 a les 16:08.
Polysphondylium pallidum
Polysphondylium pallidum és un llim cel·lular que té forma d'arbre ramificat a diferència d'altres llims com Dictyostelium discoideum que tenen una morfologia no ramificada. Principalment viu en el terra dels boscos o en els fems. La seva distribució és força àmplia i es pot trobar a Àfrica, a América, al Canadà i al Japó.La mitosis és un procès molt estudiat en P.pallidum i s'ha determinat que el seu nombre de cromosomes és 7.
| Regne | Eukaryota |
| Fílum | Amoebozoa |
| Classe | Mycetozoa |
| Ordre | Dictyosteliida |
| Família | Dictyosteliaceae |
| Gènere | Polysphondylium |
| Espècie | P. pallidum |

Per a més informació podeu veure l'entrada de Viquipèdia per a Polysphondylium pallidum que hem creat l'11 març 2011 a les 16:17.
Pythium ultimum
Pythium ultimum és un patogen que es troba en el sòl i infecta a plantes, per això es considera que causa grans pèrdues econòmiques a nivell mundial. Aquest té una amplia distribució terriorial així com també un rang d’hostes molt extès.| Regne | Chromalveolata |
| Fílum | Heterokontophyta |
| Classe | Oomycetes |
| Ordre | Pythiales |
| Família | Pythiaceae |
| Gènere | Pythium |
| Espècie | P.ultimum |
Per a més informació podeu veure l'entrada de Viquipèdia per a Pythium ultimum que hem creat l'11 març 2011 a les 16:38.
Saprolegnia parasitica
Saprolegnia parasitica és un oomicet patogen que ataca a diverses varietats de peixos, amfibis i crustacis importants per mantenir l’ecosistema aquàtic. A més a més, aquest patogen causa pèrdues econòmiques molt importants a nivell mundial, ja que el salmó, el bagre i la truita són susceptibles a ser infectats per aquest.| Regne | Protist |
| Fílum | Heterokonta |
| Classe | Oomycota |
| Ordre | Saprolegniales |
| Família | Saprolegniaceae |
| Gènere | Saprolegnia |
| Espècie | S.parasitica |
Per a més informació podeu veure l'entrada de Viquipèdia per a Saprolegnia parasitica que hem creat l'11 març 2011 a les 13:38.
Thecamonas trahens
Thecamonas trahens és un protista eucariota unicel·lular, biflagelat i extès territorialment, tot i que es troba principalment en ambients marins.
| Regne | Protist |
| Fílum | Apusozoa |
| Classe | Thecomonadea |
| Ordre | Apusomonadida |
| Família | Apusomonadidae |
| Gènere | Thecamonas |
| Espècie | T. trahens |
Per a més informació podeu veure l'entrada de Viquipèdia per a Thecamonas trahens que hem creat l'11 març 2011 a les 16:42.
Arbre filogenètic
A continuació, adjuntem l'arbre filogenètic on estan tots els protistes que nosaltres hem analitzat:
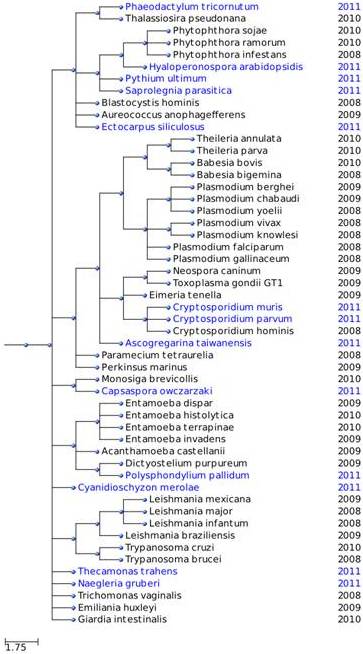
Selenoproteïnes
Les selenoproteïnes són una classe de proteïnes que contenen selenocisteïna (Sec), un aminoàcid atípic anàleg a la cisteïna, però que es diferencia d’aquesta per la presència d’un àtom de seleni enlloc de l’àtom de sofre, característic de la cisteïna.
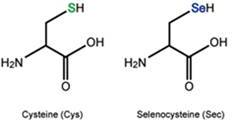
En la major part de les espècies existeixen 21 aminoàcids diferents que permeten sintetitzar les proteïnes. Generalment es parla de 20 aminoàcids, l'aminoàcid 21 és la selenocisteïna, representat per la lletra U (Sec-U).
La majoria de selenoproteïnes són enzims que participen en reaccions redox, la regió activa dels quals freqüentment manté la disposició UXXC o CXXU.
Les selenoproteïnes estan presents en els tres dominis de la vida: Bacteria, Archea i Eukarya. Els eucariotes tenen una distribució molt diferent de selenoproteïnes o selenoproteomes, que pot variar des de zero, en plantes, fins a més de trenta en alguns peixos i algues.
Actualment es coneixen 25 famílies de selenoproteïnes en eucariotes, tot i que segurament n’existeixen més. No obstant, el nombre de famílies de selenoproteïnes existents varia depenent de l’espècie. Per exemple, en Drosophila melanogaster hi ha 3 selenoproteïnes (SelK, SelH i SPS2), mentre que en Caenorhabditis elegans només n’hi ha 1 (Tioredoxina, també coneguda com TK).
Existeix certa variabilitat de selenoproteïnes a nivell interespecífic i, fins i tot, existeixen evidències de variacions intraespecífiques. És a dir, s’han descrit gens que, en comptes de codificar per selenocisteïna, tenen un gen homòleg amb una cisteïna. Un exemple d’aquest fet rau en el genoma de fugu on trobem una selenoproteïna, mentre que en el gen homòleg humà hi trobem una cisteïna.
A més a més, alguns organismes han perdut la maquinària necessària per la síntesis de selenoproteïnes durant l'evolució. Així doncs, els gens d’aquests organismes, en comptes de codificar per selenocisteïna tenen un gen homòleg amb una cisteïna (Cys).

El seleni (Se) és un element traça, poc abundant a la Terra, i essencial per la vida. Aquest micronutrient està implicat en processos redox, motiu pel qual se li assigna una possible funció antioxidant. No obstant, el paper del seleni sobre els individus encara està en vies d’estudi. Ara bé, és un fet que tant l’excés com el dèficit de seleni té efectes perjudicials en la salut humana.
El dèficit de seleni dóna lloc a la coneguda malaltia de Keshan o bé a Kashin-Beck o infertilitat masculina, mentre que el seu excés és tòxic ja que no es pot metabolitzar adequadament i dóna lloc a selenosi.
D’altra banda, estudis epidemiològics indiquen que suplements de seleni disminueixen la incidència de determinades malalties neurodegeneratives, així com també és un agent quimioprotector del càncer, redueix l’expressió viral i prevé de problemes cardiovasculars i trastorns musculars.
Com veiem, la història de les selenoproteïnes és molt complicada, ja que no es coneix cap patró que indiqui si les selenoproteïnes eren ancestrals i es van perdent o bé si van apareixent noves selenoproteïnes. Per això, els estudis actuals estan enfocats en estudiar els patrons evolutius d’aquesta classe de proteïnes tant particular i intentar respondre qüestions trivials, com ara la raó per la qual C.elegans manté tota la maquinària necessària per a sintetitzar una sola selenoproteïna.
Per a més informació podeu veure l'entrada de Viquipèdia per a Selenoproteïna que hem editat el 15 de març 2011 a les 10:09.
El codó de la selenocisteïna és UGA
El codó que defineix la selenocisteïna és el codó stop UGA. Així doncs, ens trobem davant d’un cas únic, UGA és el primer codó que té dos significats diferents dins d’una mateixa espàcie. Per aquest motiu, les selenoproteïnes estan molt mal anotades ja que la majoria de programes assumeixen que l’UGA és un codó stop i, per tant, les selenoproteïnes no són reconegudes.
Biosíntesis de selenoproteïnes
La síntesis de selenoproteïnes és un procés evolutivament conservat, on el tRNA de la selenocisteïna es sintetitza a partir del tRNA de la serina, gràcies a la intervenció dels següents factors:- SPS1 i SPS2 (Selenofosfat Syntase 1 i 2).
- eEFSec (Selenocysteine Especific Elongation factor)
- SBP2(SECIS Binding Protein 2) s'encarrega d'unir l'element SECIS i el ribosoma.
- tRNASec: tRNA específic per Selenocisteïna.
- Ribosomal protein L30: component ribosomal que també està implicat en la unió de l'element SECIS.
- Sec43p: forma part del complex format per SCP2/tRNAsec/eEFSec, peró la seva funciò exacta ès desconeguda.
- SecS (SLA/LP) (Eukaryotic Selenocysteine Syntase): converteix el Ser-tRNASec a Sec-tRNASec.
- PSTK (Phosphoseryl tRNA kinase): fosforil·la el Ser-tRNASec.
Per veure una descripció més acurada de cada factor podeu clicar a sobre de cada nom.
Elements SECIS
Existeix un mecanisme que permet saltar-se el codó stop i incorporar l’aminoàcid selenocisteïna en la cadena polipeptídica que s’està sintetitzant, l’element SECIS (SElenoCysteine Insertion Sequence).
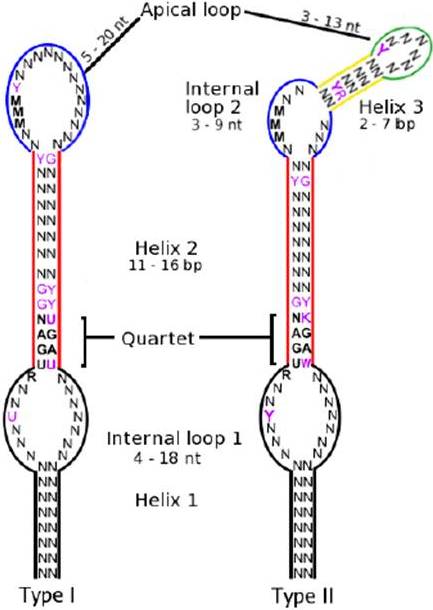
L’element SECIS és una estructura tridimensional en forma de loop i formada per 60 nucleòtids. En eucariotes i Archea, els SECIS es localitzen en l’extrem 3’ UTR dels mRNA codificants per selenoproteïnes, mentre que en bacteris es situa downstream després del codó UGA, també en posició 3’.
En el core de l’element SECIS hi ha 4 nucleòtids conservats, la funció principal dels quals és dirigir la maquinària de traducció per tal que el codó UGA sigui reconegut com a selenocisteïna i no com un codó stop.
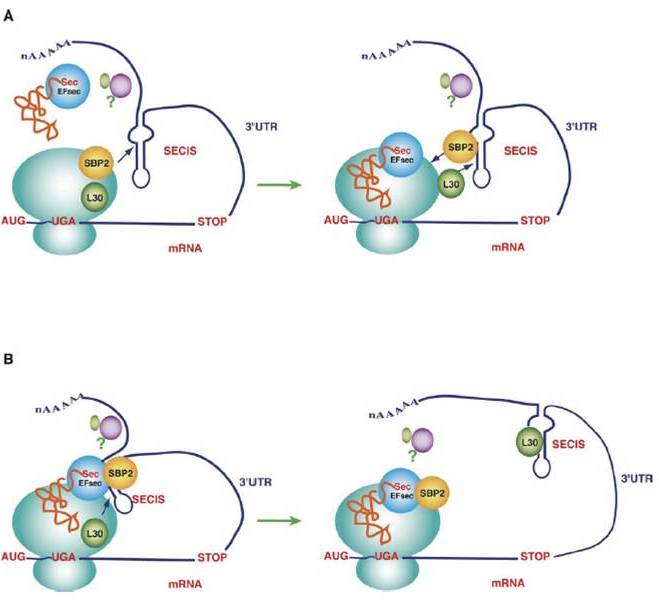
El procés de recodificació del codó UGA s’inicia quan l’element SECIS recluta la proteïna SBP2 (SECIS Binding Protein 2), la qual, al seu torn, recluta el factor d’elongació eEFSec. Aquest factor d’elongació és l’encarregat de portar el tRNA Sec específic amb l’aminoàcid selenocisteïna.
Finalment, el ribosoma, en comptes de finalitzar la traducció de la cadena polipeptídica perquà ha trobat un codó stop UGA, reconeix l’UGA com una selenocisteïna i continua la lectura de la cadena polipeptídica fins al següent codó stop.
La base de dades SelenoDB conté algunes selenoproteïnes.
El nostre treball està enfocat en la cerca de tres famílies de selenoproteïnes, la EhSEP2, la SelO i la SelS.
Familíes de Selenoproteïnes estudiades
EhSEP2
S’han reportat sis proteïnes que contenen seleni en Emiliania huxley, una alga dependent de seleni pel seu creixement. E. huxley té una nova via metabòlica que actua en la síntesi de selenoproteïnes i, com a tal tindrà nous tipus de selenoproteïnes i un sistema d’inserció de Sec diferent.
Es va identificar una nova selenoproteïna eucariota en E.huxley, EhSEP2. EhSEP2 és una proteïna de 27 kDa homòloga a la Proteïna Disulfite Isomerasa (PDI) i conté un domini tiorredoxina molt conservat. Aquesta està codificada per TGA però, a priori, no es va identificar cap SECIS.
L’estructura SECIS-like d’EhSEP2 s’assembla més a un element SECIS arcaic que a un eucariòtic per l’alt contingut en GC de les hèlix, les quals tindran una estructura rígida. EhSEP2 té un motiu KDEV al C terminal, que és molt semblant a la senyal de retenció en el reticle endoplasmàtic de mamífers (KDEL); això suggereix que EhSEP2 es localitza en el RE, igual que PDI. A més a més, es troba glicosilada com moltes altres PDI.
Per tant, si EhSEP2 funciona com una PDI localitzada al RE, una deficiència de seleni resultaria en una disfunció d’aquesta proteïna, causant un retràs en la maduració de les noves proteïnes sintetitzades i s’acabaria aturant la divisió cel·lular. Aquesta és una resposta raonable a perquè un organisme fotosintàtic, com E.huxley, necessita seleni.
Les proteïnes semblants a PDI solen tenir 1, 2 o 3 dominis TRX i un motiu CGHC. EhSEP2 només conté un domini TRX i, en EhSEP2, la Sec es localitza en el motiu UGHC a la posició corresponent al lloc actiu de les proteïnes PDI-like. Com que l’activitat de PDI és dependent de la reactivitat de la cisteïna N-terminal, es pot pensar que la substitució del residu cisteïna per una selenocisteïna en el lloc actiu de EhSEP2 és per augmentar l’eficiència catalítica d’aquesta PDI amb només un domini.
Sel O
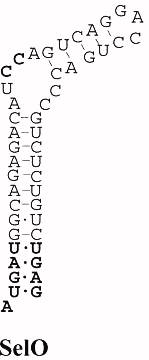
La família SelO té homòlegs en animals, bactèries, llevats i plantes però la seva funció encara és desconeguda. Les úniques proteïnes homològues a Sel O que tenen selenocisteïna són les de vertebrats i tenen l’aminoàcid localitzat en la regió C-terminal. En humans, aquesta proteïna té una llargada de 669 aminoàcids.
En altres organismes, com en S.cerevisiae, es tracta d’una proteïna homòloga que té reemplaçada la selenocisteïna per una cisteïna i, conseqüentment, la llargada de la proteïna és de 688 aminoàcids.
Aquesta proteïna està caracteritzada per tenir SECIS atípics, en els que les citosines reemplaçen les adenosines conservades del loop apical. A més a més, conté un motiu CXXU que suggereix que aquesta proteïna podria tenir una funció redox.
Per a més informació podeu veure l'entrada de Viquipèdia per a Sel O que hem creat l'11 març 2011 a les 18:57.

Sel S
La família SelS està formada per 189 aminoàcids en humans i 190 en porcs, tenint un pes molecular aproximat de 22kDa. En el genoma humà, el gen que la codifica s’anomena SEPS1 i es troba en el cromosoma 15 a la regió q26.3.
La SelS humana és una proteïna de membrana del reticle endoplasmàtic, que participa en la retrotranslocació de les proteïnes mal plegades cap al citosol, on seran degradades. En situacions d’estrés, l’expressió del gen SEPS1 es veu up-regulada, per tal d’evitar l’acumulació de proteïnes mal plegades, sovint relacionat amb procesos inflamatoris.
A més a més, l’up-regulació d’aquest gen també s’ha relacionat amb la privació de glucosa.
Així doncs, es conclou que SelS es troba associada a la inflamació, susceptibilitat al càncer colonorectal, malalties coronàries, artritis, diabetis, etc.