|
 |
| |
|
|
| |
|
|
| |
Descripció a
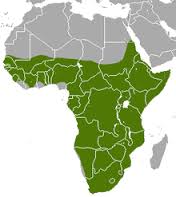
L'Orycteropusafer, porc formiguer o aardvark és un mamífer de la família Orycteropodidae que viu a pràcticament tot continent Africà, al sud del desert del Sàhar, excepte a la regió de Namibals. Viu a la sabana i a zones boscoses, és un animal solitari i nocturn, durant el dia dorm als caus i només es relaciona per a aparellar-se. S'alimenta de formigues i termites, les obté directament dels seus nius, per això tenen una llengua molt llarga i les dents especialment modificades.
|
|
| |
Pel que fa a la morfologia, té la pell molt gruixuda i no té capa de greix. La seva similitud amb els ossos formiguers americans i pangolins és simplement aparent i degut a convergència evolutiva. És de color cru a la part superior i vermellós a la inferior, el cos és robust i l'esquena arquejada. És un animal digitígrad amb les potes anteriors curtes, fortes i sense polze, i les posteriors amb cinc dits. Els dits tenen ungles grans amb forma de pala aplanada. [1,2]
La pell és dura i amb poc pèl. El seu pes varia entre els 40 i 65 Kg, té una longitud mitjana d'entre 1 i 1,7 metres, tot i que pot mesurar fins 2,2 metres. Els seus ulls estan envoltats de pèls amb funció sensorial i són daltònics.
L'esperança de vida mitjana de l'espècie en llibertat és d'uns 18 anys, però en captivitat viuen al voltant de 23 anys. [2]
|
|
|
|
| |
El seu genoma presenta només deu cromosomes, caràcter primitiu que s'ha perdut en els euteris més avançats. L'Orycteropus afer és l'única espècie vivent del ordre Tubulidentata. [1]
Tornar a dalt |
|
| |
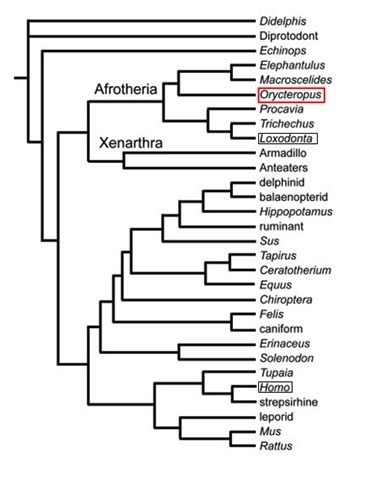
Taxonomia i arbre filogenètic a
Segons el NCBI el seu llinatge és: Cellular organisms; Eukaryota; Opisthokonta; Metazoa; Eumetazoa; Bilateria; Deuterostomia; Chordata; Craniata; Vertebrata; Gnathostomata; Teleostomi; Euteleostomi; Sarcopterygii; Dipnotetrapodomorpha; Tetrapoda; Amniota; Mammalia; Theria; Eutheria; Afrotheria; Tubulidentata; Orycteropodidae; Orycteropus.
Tornar a dalt |
|
| |
|
|
| |
|
|
| |
El seleni a la natura a
El seleni és un micronutrient essencial per a animals, microorganismes i altres eucariotes. La seva importància s’ha atribuït principalment a les Selenoproteïnes de les quals en forma part a través del’aminoàcid selenocisteina (Sec). [4]
Tant el dèficit com l'excés d'aquest micronutrient comporta efectes perjudicials per a la salut. El dèficit de Seleni, per exemple, es pot traduir en la malaltia de Keshan, caracteritzada per una necrosis a nivell de miocardi que comporta una disfunció cardíaca. A més a més, la suplementació amb seleni s'ha correlacionat amb la prevenció d'alguns càncers humans. [5,6]
|
|
| |
Les Selenoproteïnesa
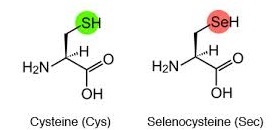
Les selenoproteïnes es caracteritzen per tenir una selenocisteïna en la seva estructura. La selenocisteïna és l'aminoàcid número 21 i es representa amb les abreviatures U o Sec. Aquest aminoàcid té una estructura molt similar a la cisteïna però incorpora un àtom de seleni en comptes de sofre.
El seleni i el sofre tenen propietats químiques molt semblants, i degut a això, en la recerca de gens ortòlegs que codifiquen per selenoproteïnes, moltes vegades es troben cisteïnes en el seu lloc.
En els organismes, el seleni es troba formant part de les selenoproteïnes, i no lliure dins les cèl·lules, permetent així mantenir-lo com a element essencial però evitant la seva toxicitat. La majoria de les selenoproteïnes són enzims que participen en reaccions d’oxidació-reducció, essent la selenocisteïna un residu essencial del centre catalític, i tenint així una funció antioxidant. A més, també tenen funció antitumoral, immunològica i participen en el metabolisme d’hormones tiroidees. [6,7]
Un aspecte a considerar és que es pot reduir el selenoproteoma a través de la conversió de Sec a Cys. Aquest procès és específic de les selenoproteïnes i pot succeir degut a una mutació puntual que transformi el codó UGA de la Sec en un codó que codifiqui per a Cys. No obstant, la Sec i la Cys no són funcionalment equivalents, per això, les conversions a Cys no són neutrals. [8]
|
|
|
|
|
| |
Síntesia
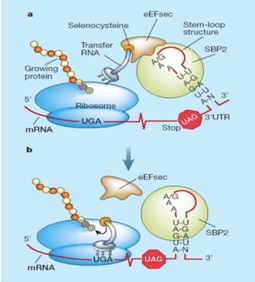
El codó que s’assigna a les selenocisteïna és el codó UGA, i normalment aquest s’interpreta com un STOP que al ser reconegut per la maquinària traduccional fa que el procés de síntesi s’acabi. Però en el cas de les selenoproteïnes aquest codó no s'interpreta com un STOP sinó que codifica per una Selenocisteina i continua la traducció.
En eucariotes, el mecanisme mitjançant el qual aquest codó s’interpreta de manera diferent esta mediat per l’extrem 3’ de la regió UTR al final del mRNA. Els gens que codifiquen per selenoproteïnes contenen en aquest extrem una estructura tridimensional en forma de loop anomenada SECIS (Selenocystein Insercion Sequence), la qual reclutarà la maquinària necessària per a sintetitzar una selenoproteina. [7]
Primer, cal la intervenció de SBP2 ( SECIS Binding Protein), la qual s'uneix a SECIS i reclutarà un factor d’elongació específic eEFsec (SEC elongation factor), el qual aproxima el tRNA de la selenocisteïna (tRNAsec) al codó UGA del mRNA de les selenoproteïnes i determina la traducció a selenocisteïna. La traducció del mRNA segueix fins el reconeixement d’un nou codó de terminació.
A més dels components especificats anteriorment, per a la síntesi de l’aminoàcid Sec hi ha involucrats aproximadament unes 10 proteïnes o factors que són necessaris per tal que un genoma pugui sintetitzar selenocisteïna:
- tRNAsec → és el tRNA encarregat d’incorporar la selenocisteïna en la cadena polipeptídica.
-
SPS1 i 2 (selenofosfat sintetases 1 i 2)→ sintetitzen selenofosfat a partir del seleni.
-
PSTK (fosfoseril tRNA quinasa) →fosforila el seril-tRNA a O-fosfoseril-tRNA, que és un intermediari actiu per a la síntesi de selenocisteïna.
-
SLA/LP o SecS (selenocisteïna sintasa)→converteix O-fosfoseril-tRNA en selenocistenil-tRNA, necessari per a la síntesi de selenoproteïnes.
-
SECp43 (proteïna associada a selenocisteïna 1)→ Té un paper important en els primers passos de la síntesi de selenocisteïna i en la càrrega del tRNA corresponent que permetrà la incorporació de la selenocisteïna a les selenoproteïnes. Estabilitza el complex SBP2 - eEFSec - tRNA afavorint l’eficiència de la síntesi de selenoproteïnes. [9,10,11]
|
|
| |
Tornar a dalt |
|
| |
Predicció de selenoproteïnesa
Cal considerar que la identificació automatitzada de les selenoproteïnes es veu limitada per la consideració sistemàtica que fan els programes informàtics dels codons UGA com a stop-codons i no com a triplets codificant per a una selenocisteïna. Val a dir que és només en un 25 dels casos on això no es compleix perquè es tracta d’una selenoproteïna, així que l’error és petit, però hi és. A primera vista, un bon mètode per discriminar entre aquestes dues "lectures" del mateix codó podria ser buscar els SECIS, si no fos perquè aquests no presenten un patró conservat de la seqüència. Tot i així, es coneix que hi ha SECIS que conserven el quartet AUGA i altres que contenen altres quartets, anomenats canònics i no canònics respectivament. Així doncs, l'estratègia òptima per predir la presència de selenoproteïnes en un genoma, es basa tant en la informació sobre els exons del gen d'estudi com en la identificació dels elements SECIS. |
|
| |
Tornar a dalt |
|
|
|
|
|
|
|
|
|
|
|
|
|
|