|
Família DI
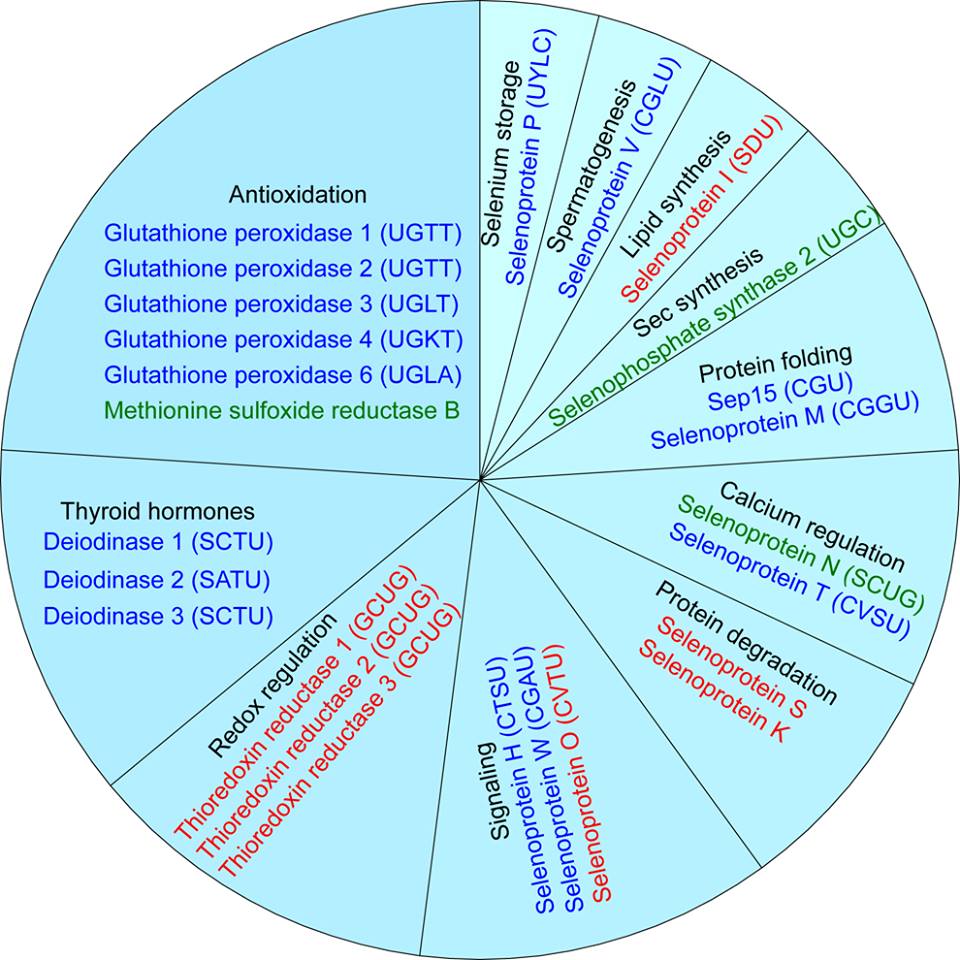
La família DI (Iodotironina Deiodinases), o DIO, és la responsable de connectar la biologia del seleni amb el metabolisme de les hormones tiroidees. Aquesta família, en mamífers, consta de tres oxidoreductases amb selenocisteïna (Sec) distribuïdes de manera ben diferenciada: DI1, DI2 i DI3. Aquests tres enzims catalitzen l'activació (DI1 i DI2) i la inactivació (DI3) de les hormones tiroidees T4 (tiroxina), T3 (triiodotironina) i rT3 (reverse-triiodotironina) mitjançant l'eliminació de diferents restes de iode.
Aquestes hormones regulen el creixement, l’audició i varis processos metabòlics, com la termogènesi. A més, són indispensables pel desenvolupament normal del cervell fetal i controlen l'expressió gènica en tots els teixits vertebrats via receptor d'hormones tiroidees (TR).
S'ha vist que la DI1 pot jugar un paper més important en la producció de T3 a la glàndula tiroide així com controlar el nivell de T3 circulant, mentre que la DI2 i DI3 es pensa que tenen una funció local en els processos de deiodinació, a nivell d'òrgan o teixit específic.
A part de diferències en la seqüència d'aminoàcids, totes les DIs són estructuralment semblants: són proteïnes de membrana que adopten una estructura tioredoxin-foldlike amb una Sec present al centre actiu, en concret es troba en un motiu molt característic compost per: FGSXTU. La DI1 i DI3 es localitzen a la membrana plasmàtica mentre que la DI2 resideix en la membrana del RE. [12]
En el nostre anàlisi de l'Orycteropus afer s'han trobat les tres selenoproteïnes, tant utilitzant la query humana com la de l'elefant, és a dir els tres membres de la família de selenoproteïnes humanes DI es conserven en Orycteropus afer. A més en les tres s'han trobat elements SECIs, fet que ens confirma la seva existència:
DI1
En referència a la selenoproteïna humana, el percentatge d'homologia més alt es troba en l'scaffold gi|406064300|gb|JH863863.1| (86,76%) ara bé, aquest no és el hit amb l'e-value més alt, sinó el segon de la llista (3e-27) però com que en l'anàlisi de la selenoproteïna en elefant ens dóna el mateix scaffold, i en aquest cas si que és l'e-value més gran, el prenem com a millor hit i per tant, com a regió que contindrà la selenoproteïna. Descartem els altres scaffolds i considerem que la seqüència obtinguda de l'scaffold esmentat correspon a la selenoproteïna que estem estudiant (DI1). A més, es pot confirmar gràcies a trobar el motif -SCTU- en la seva seqüència.
DI2
En referència a la selenoproteïna humana, el percentatge d'homologia més alt es troba en l'scaffold gi|406065015|gb|JH863797.1| (veiem un hit en aquest de 94% d'homologia amb la query). A més en aquest scaffold també és on hi ha el hit amb l'e-value més alt (6e-97). Al fer l'estudi amb la selenoproteïna de l'elefant com a referència també ens surt aquest scaffold com a regió que conté el millor hit. En ambdós casos trobem la selenocisteïna ben alineada i per tant podem dir que la seqüència obtinguda de l'scaffold esmentat correspon a la selenoproteïna que estem estudiant (DI2). A més, també hi trobem el motiu del centre catalític típic d’aquesta selenoproteïna -SATU-.
DI3
En aquest cas, la selenoproteïna humana que utilitzem com a model ens dóna un alineament al blast que ens permet discriminar molt fàcilment l'scaffold que hem d'utilitzar: gi|406062479|gb|JH864089.1|. Doncs el hit que es troba en aquest scaffold té un boníssim e-value (1e-146) i un boníssim grau d'homologia (94%). Al fer l'estudi amb la selenoproteïna de l'elefant com a referència també ens surt aquest scaffold com a regió que conté el millor hit. En ambdós casos trobem la selenocisteïna ben alineada i per tant podem dir que la seqüència obtinguda de l'scaffold esmentat correspon a la selenoproteïna que estem estudiant (DI3). En aquest cas tornem a trobar el motiu esperat: -SCTU-.
Pel que fa a la cerca de SECIs realitzada al SeciSearch3, s’han trobat varis elements: 4 per DI1, 1 per DI2 i 2 per DI3. Això ens confirma el fet d’haver trobat les selenoproteïnes.
Aquests SECIs també els trobem en els homòlegs en Cys d’aquesta família (DI1 i DI2). Això no indica que aquestes seqüències dels homòlegs continguin selenoproteïna, el fet de trobar elements SECIs es pot explicar degut a la coincidència d’scaffolds que veiem: DI1 i DI2 comparteixen els mateixos scaffolds que les dues selenoproteïnes amb el mateix nom. Ara bé, no pertanyen a la mateix regió, per això considerem que són dues proteïnes diferents, però com que amb el programa de cerca de SECIs agafem una llargada de la seqüència upstream de 5000 nucleòtids, segurament ens trobem que englobem la regió de les selenoproteïnes i en detectem els seus SECis. De fet, són seqüències molt semblants però s’acaben abans de la regió on trobem la U en les selenoproteïnes.
Tornar a dalt
Família GPx
Les glutatió peroxidases (Gpx) pertanyen a una família d’enzims relacionats filogenèticament. Aquestes realitzen la funció de defensa enzimàtica antioxidant ja que catalitzen la reducció del peròxid d'hidrogen i hidroperòxids orgànics protegint les cèl·lules del dany oxidatiu. A més, modulen la síntesi d'eicosanoides, regulant la inflamació, i disminueixen el dany oxidatiu de lípids, lipoproteïnes i DNA.
En mamífers GPX1-4 són selenoproteïnes amb una Sec al centre catalític, i GPx6 és una selenoproteïna únicament en algunes espècies. D’acord amb la filogenia, la família GPx conté tres grups evolutius derivats d’un ancestre amb Cys: GPx1/GPx2, GPx3/GPx5/GPx6 i GPx4/GPX7/GPx8. Les proteïnes GPx7 i GPx8 que són homologues en cisteïna evolucionen d’un antecessor de GPx4; GPx5 i GPx6 apareixen com a resultat d’una duplicació en tàndem de GPx3.
Aquests enzims difereixen en l'especificitat pel substrat de la degradació de peròxids i en la distribució tissular, la seva localització específica és la següent:
-
GPx1 citosol cel·lular o gàstrica
-
GPx2 gastrointestinal
-
GPx3 plasmàtica o extracel·lular
-
GPx4 isoformes citosolica, nuclear i mitocondrial
-
GPx6 epiteli olfactori i teixits embrionaris
Homolègs de cisteïna:
GPx1
En la comparació amb les selenoproteïnes humanes primerament hem comparat amb l’scafflold gi|406066660|gbJH863738.1 perquè és el que té un e-value més petit (3e-72), tot i que el seu grau d'homologia (75,53%) no és el més elevat. Hem seguit el mateix criteri amb la comparació amb les selenoproteïnes d’elefant, essent l’scaffold amb el millor e-value el mateix, amb un percentatge d’homologia de 75,98%. Podem dir que la selenocisteïna es conserva ja que se'ns alinea en els dos casos una U de la query amb una U del nostre scaffold.
Però si utilitzem l’scaffold gi|406062107|gb|JH864216.1 que presenta la homologia més elevada (95,24%) en Loxodonta i en humans (94,37%), i amb un e-value molt petit també (4e -27 en Loxodonta i 7e -31 en humans) també observem una U alineada amb una U. Per tant, trobem una duplicació d’aquesta selenoproteïna en l’Orycteropus afer perquè està present en dos scaffolds diferents. Deduïm que això és degut a la presencia d’un pseudogen d’aquesta ja que en la bibliografia consultada hem vist que en mamífers tenen un pseudogen localitzat al cromosoma X, però no tenim suficient informació per a confirmar-ho.[8]
Per comprovar l’existència de la selenoproteïna hem mirat la presència de SECIs mitjançant el programa SeciSearch. Aquest ens detecta un SECI en els dos scaffolds.
Finalment concloem que és la proteïna en qüestió perquè trobem el motiu carecterístic d’aquesta, -UGTT-, present en el centre catalític.
GPx2
En aquest cas, al comparar amb la selenoproteïna humana ens dóna un alineament al blast que ens permet seleccionar l’scaffold fàcilment, aquest serà el gi|406064883|gb|JH863806.1, ja que el hit d’aquest té el grau d’homologia més elevat (95%) i el millor e- value (4e -62). En utilitzar com a model la selenoproteïna de Loxodonta també ens dóna aquest scaffold com a regió que conté el millor hit. Al mirar l’alineament d’aquest observem en els dos casos que la selenocisteïna està ben alineada, i a més, presenta el motiu esperat del centre catalític, -UGTT-. Per tant, podem predir que la seqüencia de l’scaffold que hem seleccionat correspon a la selenoproteïna GPx2. Posteriorment hem analitzat la presència de SECIs en aquest scaffold i el resultat és positiu.
GPx3
És la selenoproteïna més abundant en el plasma sanguini, on es secreta de forma glicosilada.
En aquesta, a l’hora de seleccionar l’scaffold, ens trobem amb el mateix cas que en l’anterior, hem seleccionat el gi|406064636|gb|JH863828.1, ja que presenta el major grau d’homolgia (97%) i el millor e-value (2e -31) tant si comparem amb humans com en Loxodonta. Al comparar el nostre hit amb les dues seqüències model observem que la Sec, en el motiu -UGLT-, es conserva. Per això, afirmem que la seqüència d’aquest scaffold correspon a la selenoproteïna GPx3. A més, s’han trobat SECIs en aquesta regió, fet que confirma la predicció.
GPx4
És un enzim que actua en l'oxidació dels lípids i és essencial per a la vida, ja que té un rol essencial en la diferenciació cel·lular durant el desenvolupament embrionari, maduració de l'esperma i fertilitat dels mascles.
Al comparar amb la selenoproteïna humana escollim l’scaffold gi|406062139|gb|JH864184.1 perquè és el que presenta el grau d’homologia més elevat (92,1%) i l’e-value més petit (2e -22). Si utilitzem com a model la d’elefant observem que el mateix scaffold també és el que presenta més homologia (95,9%) i un e-value millor (2e-20). Al alinear les dues seqüències models amb el nostre hit observem que la Sec està conservada i a més, presenta el motiu característic d’aquesta, -UGKT-, per tant, concloem que la seqüència d’aquest scaffold correspon a la selenoproteïna GPx4. Hem detectat SECIs en aquesta regió
GPx5
A diferència de les glutatió peroxidases que s’han comentat anteriorment, aquest mRNA no conté el codó de selenocisteina (UGA) i per tant, la proteïna codificada és independent del seleni. S’ha proposat que jugui un rol en la protecció d’efectes nocius de la peroxidació lipídica i/o prevenint de la reacció acrosòmica prematura en la membrana dels espermatozoides.
Quan utilitzem la query de l’elefant, veiem que la Cys s’alinia en el genoma de l’Orycteropus afer amb una Cys, podem dir que conservem la GPx5. A més, es veu en la seqüència comparada amb Loxodonta el motiu del centre catalític:- NVATYCG-.
Ara bé, en quant a la query de l’humà s’alinia molt malament i no aconseguim trobar l’homòleg en cistcisteïna estudiat.
Com que hem trobat un element SECI per a aquesta proteïna hem sospitat que podria ser que en el nostre organisme hi hagués una selenocisteïna i a més, utilitzant el programa Seblastian ens prediu una selenoproteïna, concretament ens dóna un alineament amb el precursor de GPx6 de Sus scrofa (porc senglar). Per aquest motiu, hem considerat oportú realitzar una altra vegada l’anàlisi utilitzant com a query la GPx6 de S.scrofa, obtinguda del Gene Script. Els resultats obtinguts no indiquen presència de selenocisteïna, ara bé, aconseguim un millor alineament que en humans i podem concloure que és un homòleg de Cys ja que trobem el motiu -NVATYCG-. Consultant la bibliografia [8] s’ha vist que la GPx5, en espècies que són més antigues evolutivament tenien una selenocisteïna, que a partir del Platypus (ornitorrinc) s’ha perdut donant lloc a una Cys, com és el nostre cas.
GPx6
En aquest cas hem seleccionat l’scaffold gi|406061978|gb|JH864345.1 ja que, en la comparació amb humans té el grau d’homologia més elevat (85%) amb un e-value de 7e-30, i en Loxodonta presenta un grau d’homologia del 100% amb un e-value de 1e-32. A l’alinear les seqüències trobem tant en el cas de la query d'humans com en Loxodonta que la U s'alinea amb una C del nostre scaffold, per tant, en l'O.afer tenim un homòleg de cisteïna enlloc d'una selenoproteïna. Per això tampoc conserva el motiu característic del centre catalític d’aquestes selenoproteïnes -UGLA-, sinó que hi trobem -CGLT-. Aquesta conversió a Cys, només es troba en algunes espècies concretes dins la classe dels mamífers. No obstant, observem que encara hi ha conservat un element SECI en aquest scaffold.[8]
En el cas de GPx5, GPx7 i GPx8 que són proteïnes homòlogues en cisteïna, (CysGPxs) el centre actiu es troba, com els SecGPxs, en un motiu NVAxxC(Sec)G aprop de l’N-terminal. [13]
GPx7
És una glutatió peroxidasa monomèrica del RE que també conté una Cys en el seu centre redox.
Hem seleccionat l'scaffold gi|406060746|gb|JH865577.1| en humans i Loxodonta. Al comparar els resultats amb la query humana i la de Loxodonta veiem que s'alinea una C amb una C del nostre scaffold, per tant, l'Orycteropus afer conserva l'homòleg de cisteïna de la GPx7. A més, veiem el motiu propi en la seva seqüència del centre actiu: -NVASECG-
GPx8
En aquest cas també veiem que es conseva la Cys, tant en l’alineament amb humans com en loxodonta, que conté el seu centre redox. (-NVASDCQ-).
Tornar a dalt
Família TR
Les proteïnes TrxR (conegudes com TRs) formen part, juntament amb la thioredoxina (Trx) i NADPH, del sistema thioredoxina, que és el major sistema cel·lular redox present en organismes vius. És un sistema indispensable per al desenvolupament i probablement per a la proliferació cel·lular in vivo.
Les TrxRs contenen diferents dominis: domini FAD (CxxxxC), domini d'unió a NADPH, domini d'interfase i un residu Sec (Selenocisteïna) en l'extrem C-terminal, en la penúltima posició de la seqüència, que és indispensable per a la seva activitat enzimàtica. Aquests enzims són homòlegs a la glutatió reductasa, ja que el domini catalític C-terminal de l'enzim és idèntic en les dues proteïnes (GCUG).
Hi ha dues formes d'aquestes proteïnes TrxR: una present en bacteris i una altra present en eucariotes superiors, com mamífers. Les TrxRs de mamífers tenen un paper important en molts processos cel·lulars, com: reducció de proteïnes encarregades de processos molt importants per a l'organisme, regulació de l'expressió gènica de diversos factors de transcripció, regulació indirecta de la proliferació cel·lular, la mort cel·lular i l'activació de la resposta immune, també participen en el metabolisme del Seleni i controlen la síntesi de selenoproteïnes.
Hi ha 3 selenoemzims TrxR descrits en mamífers:
-
TrxR1 --> enzim citosòlic. S'ha vist que la seva deleció condueix a la mort de l'embrió degut a un gran retard del creixement i un desenvolupament anormal de molts teixits, inclòs el cor.
-
TrxR2 --> enzim mitocondrial. Essencial per a l'hematopoesi i el desenvolupament cardíac, altrament dit, si no hi ha TrxR2 es produeix la mort de l’embrió.
-
TrxR3 --> enzim específic dels testicles
També s'han descrit splicings alternatius que suggereixen l'existència d'altres espècies de TrxRs.
En els nostres anàlisis, utilitzant les querys humanes hem aconseguit identificar TR1 i TR2 en l'Orycteropus afer, però no TR3.
TR1
La TR1 es troba en l'scaffold gi|406062751|gb|JH864052.1| amb una homologia del 98,7% i un e-value de 5e-35. En aquest mateix scaffold, hi trobem una estructura SECIS de grau A, fet que ens confirma que es tracta d'una selenoproteïna. En Loxodonta no es troba aquesta forma.
TR2
La TR2 es troba en l'scaffold gi|406061776|gb|JH864547.1| amb una homologia del 97,06% i un e-value de 3e-21. En aquest mateix scaffold, hi trobem una estructura SECIS de grau A, per tant, podem afirmar que hem trobat la selenoproteïna en l'Orycterpus afer.
Al comparar el genoma de l'Orycteropus afer amb les selenoproteines de loxodonta africana només hi hem trobat la selenoproteïna TR2. Aquesta es troba en l'scaffold gi|406062751|gb|JH864052.1|, que coincideix amb l'scaffold on es troba la TR1 d'humans. Hem comprovat que no estiguessin en la mateixa posició, fet que ens indicaria que aquestes dues proteïnes podrien ser homòlogues. No es troben en la mateixa posició, per tant, l'Orycteropus afer té dues selenoproteïnes diferents en el mateix scaffold, una corresponent a la TR1 d'humans i l'altre a la TR2 de loxodonta africana.
TR3
En l’anàlisi d’aquesta, tant amb la query humana com d’elefant, s’ha vist que hi ha un pèssim alineament de la proteïna i que en l’Orycteropus afer no hi apareix la Sec alineada. Per la descripció de les selenoproteïnes, seria viable que el nostre organisme d'estudi no tingués TR3, ja que aquesta és específica de testicles i no de cap òrgan vital per a l'organisme. Passa el mateix en el cas de Loxodonta.
Pel que fa als homòlegs en cisteïna, en el cas dels humans no en trobem cap, ja que no hi ha cap seqüència d’homòlegs en cisteïna anotada en la base de dades SelenoDB. Pel que fa a Loxodonta, trobem un homòleg en cisteïna de TR en l’scaffold gi|406062751|gb|JH864052.1|, amb una homologia del 74.19% i un e-value de 5e-6. Tal com esperaríem, no trobem cap element SECIS, ja que no es tracta d’una selenoproteïna.
Sel15
Sel15, o Sep15, va ser la primera selenoproteïna identificada i té un pes de 15 kDa, d'aquí el seu nom. En el seu centre catalític s’hi troba la seqüència -CGU-. S'expressa en molts teixits, com: cervell, pulmó, testicles, fetge, glàndula tiroides i ronyons. La seva expressió es troba regulada en resposta al seleni que incorporem de la dieta. La funció exacta d'aquesta proteïna és desconeguda, però s'ha proposat que pot tenir un paper important en el plegament de les proteïnes i en la formació d'enllaços disulfur en el reticle endoplasmàtic. També podria estar implicada en la regulació de l'apoptosi i tenir importància en la supressió de tumors.
En els nostres anàlisis, tant utilitzant la query humana com la d’elefant, hem trobat que l'Orycteropus afer té la Sel15 localitzada a l’scaffold gi|406064247|gb|JH863870.1, amb una homologia del 97,67% i amb un e-value corresponent a 2e-16. En aquesta mateixa regió hi trobem un element SECIS de grau A, fet que confirma la presència d’una selenoproteïna. A més, s’ha identificat el motiu característic del seu centre catalític.
Pel que fa als homòlegs en cisteïna, en la posició on hi ha la Selenocisteïna en la Sel15, hi trobem una cisteïna en el cas de la Sel15 humana i una glicina en el cas de la Sel15 d’O.afer. Aquest fet ens indica que Sel15 en l’organisme estudiat és un homòleg en glicina i no en cisteïna. Com esperaríem, al no tractar-se d’una selenoproteïna no trobem cap element SECIS en la regió.
SelI
Aquesta selenoproteïna forma part de la família CDP-alcohol fosfatidil transferasa 1 que catalitza la síntesi de fosfatidiletanolamina a partir de CDP-etanolamina. Té un paper important en la formació i manteniment de les membranes vesiculars, és a dir, en la síntesi de lípids. Es troba expressada en molts teixits i és abundant en cervell, placenta, fetge i pàncrees. En el seu centre catalític hi trobem la seqüència -SDU-.
Segons els resultats obtinguts, podem afirmar que l'Orycteropus afer conté aquesta Selenoproteïna, tant si la comparem amb la SelI humana com amb la de Loxodonta. La SelI es troba en l'scaffold gi|406065367|gb|JH863772.1. En el cas de la SelI humana, trobem una homologia del 100% i amb un e-value de 6e-34. Per altra banda, amb al SelI de loxodonta té una homologia del 97,22% amb un e-value de 2e-12. A més, trobem la seqüència característica d’aquesta selenoproteïna.
En totes dues proteïnes, trobem un element SECIS de grau A en la mateixa regió.
SelN
Aquesta selenoproteïna s’expressa en diferents teixits i genera una glicoproteïna de 65kDa que es localitza en la membrana del reticle endoplasmàtic i es troba implicada en la regulació del calci. En el seu centre catalític hi trobem la seqüència -SCUG-. S’han descrit diverses formes de miopaties relacionades amb mutacions en la regió 5’UTR el gen de la SelN (SEPN1) i també amb mutacions entre l’element SECIS. Així doncs, tot i que la funció biològica de SelN és desconeguda, es sap que té un paper vital en el teixit muscular.
Segons els nostres anàlisis, podem afirmar que Orycteropus afer té SelN. Comparant el genoma en estudi amb la SelN d’humans i de loxodonta, veiem que aquesta es troba en l’scaffold gi|406066287|gb|JH863749.1. En ambdós casos, l’homologia de la SelN de l’Orycteropus afer i els altres dos organismes és del 97.87% i l’e-value es de 4e-20 en el cas de la SelN humana i de 3e-30 en el cas de la SelN de loxodonta.
A més, en la regió on trobem la SelN trobem dos elements SECIS, un de grau A i un altre de grau B.
SelO
És una selenoproteïna que es troba en la majoria d’eucariotes i en molts bacteris. Aquesta Selenoproteïna adopta una estructura tridimensional similar a la de les quinases, és per això que es creu que SelO, tot i no tenir el residu d’Aspartat essencial per a l’activitat catalítica, podria tenir activitat fosfotransferasa i oxidoreductasa en el seu extrem C-terminal, regió on trobem la selenocisteïna. El centre catalític d’aquesta proteïna conté la seqüència CVTU. Es creu que té activitat oxidoreductasa, ja que presenta el motiu Cxx[CU], essencial per a aquesta activitat. De manera genèrica, es relaciona aquesta proteïna amb la senyalització cel·lular.[14]
Analitzant els nostres resultats, podem afirmar que Orycteropus afer té la SelO. Aquesta es troba en l’scaffold gi|406062134|gb|JH864189.1. Presenta un 93.44% d’homologia amb la SelO humana i el hit dóna un e-value de 1e-26. En el cas de la SelO de Loxodonta, l’homologia és del 91.89% i l’e-value és 3e-35. Per a confirmar que es tracti d’una selenoproteïna, hem buscat elements SECIS en aquesta regió i no n’hem trobat cap. Cal valorar que en la SelO de l’Orycteropus afer no s’hi troba el motiu -CxxU- en el centre catalític, sinó que està subtituït pel motiu -RxxU-. No ho podem afirmar, però podria ser que aquest canvi alterés la funció de l’enzim i que repercutís en la presència o no d’elements SECIS.
Pel que fa als homòlegs en cisteïna, no en trobem cap amb els humans, però sí amb Loxodonta. En el cas dels humans no es coneix cap homòleg en cisteïna (no hi ha seqüència annotada al SelenoDB) i en el cas de Loxodonta veiem que en el centre catalític de la SelO s’ha substituït la selenocisteïna per una cisteïna tant en Loxodonta com en O.afer.
Tornar a dalt
Família Rdx12
Aquesta família proteica està formada per 4 selenoproteïnes: SelH, SelT, SelV i SelW. Presenten un domini thioredoxin-like a l'extrem N-terminal, amb un motiu altament conservat en el seu centre catalític: -CXXU-, en el cas que continguin selenocisteïna, o -CXXC, en el cas que siguin homòlegs en cisteïna. La presència d'aquest domini fa que tinguin activitat redox, de manera que una de les seves principal funcions és l'acció antioxidant.
Cada proteïna exerceix la seva activitat antioxidant en diferents tipus neuronals i/o compartiments cel·lulars: SelW es troba principalment a les neurones mentre que SelV es localitza als testicles; SelT conté un domini hidrofòbic gràcies al qual s'ancora a la membrana del reticle endoplasmàtic, però encara no se'n coneix molt bé la funció i SelH té una localització nucleolar. [12][15]
Els resultats obtinguts per les proteïnes d'aquesta família no han estat gaire bons. En la SelH no s'alinea el motiu del centre catalític i per tant, no es troba Sec en cap cas, ni utilitzant la query de H. sapiens ni la de L. africana. Pel que fa a la SelT, sí que trobem la selenocisteïna del centre catalític alineada, utilitzant la query humana, però quan utilitzem la query de L. Africana no s'alinea. Així, trobem SelT en el scaffold gi|406064334|gb|JH863860.1| de O. Afer, amb un grau d'homologia del 51,55% amb la query d'humans, i amb un e-value de 3e-17. Per SelW hem pogut alinear la selenocisteïna del motiu en ambdós casos, amb un grau d'homologia del 64,06% amb la query humana i del 51,02% amb la query de l'elefant. En ambdós casos el e-value és significatiu, essent 6e-07 i 8e-10, respectivament. La proteïna SelW es localitza a l’scaffold gi|406067052|gb|JH863720.1| de O. afer.
Finalment, de SelV només hem obtingut la query dels humans, ja que segons el SelenoDB no s'ha anotat en L. africana. No s'ha aconseguit alinear la selenocisteïna del centre catalític.
S’han identificat SECIS en SelT, SelW i SelH, però no en SelV. El fet de trobar un SECI a SelH, tot i trobar tota la proteïna ben alineada excepte el centre catalític, ens fa pensar que O. afer conserva la proteïna però ha perdut la Sec, alterant possiblement la seva funció, i per això encara conserva també els SECIs.
D’aquesta família només hem trobat homòlegs en cisteïna anotats a SelenoDB per SelW i SelT. SelW és present en H.sapiens i en L.africana, però només hem trobat hits significatius utilitzant la query d’humans. En aquest cas, hem aconseguit un alineament bastant bo, amb un 96,30% d’homologia amb un e-value significatiu (6e-17). A més, es conserva el motiu -CXXC- característic d’aquesta família. Al genoma d’O.afer, l’homòleg en cisteïna de SelW es troba en l’scaffold gi|406064934|gb|JH863803.1|.
SelT només és present en H. sapiens, i hem aconseguit alinear-la amb el genoma de O. afer amb una homologia del 100% i un e-value de 3e-18. En aquest cas no existeix el motiu -CXXC-, però les dues úniques cisteïnes presents a la query alineen amb una cisteïna en la seqüència obtinguda en l’organisme estudiat, per tant podem considerar que es conserva la homologia en cisteïna. Aquesta proteïna es localitza en el scaffold gi|406065208|gb|JH863784.1.
Així doncs, hem pogut localitzar SelT i SelW en el genoma de O.afer, però no hem trobat ni SelH ni SelV.
SelS i SelK
Aquestes dues proteïnes es troben al reticle endoplasmàtic i s’encarreguen d’eliminar les proteïnes mal plegades per tal que no s’acumulin. S’ha observat que l’expressió de SelS és induïda en resposta a estrès en el reticle endoplasmàtic, normalment relacionat amb processos inflamatoris, per a evitar que s’acumulin proteïnes mal plegades al seu interior i siguin degradades al citosol. També s’ha vist que SelS interacciona amb una proteïna que es troba en la fase aguda de la resposta inflamatòria, la SAA. Per la seva banda, SelK està involucrada en el flux de calci del reticle endoplasmàtic.[16]
SelS
Analitzant els nostres resultats, podem afirmar que O.afer conté la SelS. Trobem la SelS en l’scaffold gi|406066672|gb|JH863737.1, amb una homologia del 82.61% entre la SelS de Loxodonta i la d’O. afer, amb un e-value de 6e-13. En el cas de SelS humana, l’homologia és del 65.50%, amb un e-value de 5e-14. Aquestes dades ens indiquen que aquesta selenoproteïna es troba més conservada entre O. afer i Loxodonta que amb humans. Finalment, hem comprovat la presència d’elements SECIS en aquesta regió per a confirmar que realment es tracti d’una selenoproteïna i hem trobat 3 elements SECIS, un de grau A i dos de grau B.
SelK
La SelK la trobem molt ben alineada, tant amb la query de H. sapiens com amb la de L. africana, ja que presenta una homologia del 92% amb els primers i un 80% amb els segons. Es troba en el scaffold gi|406065809|gb|JH863759.1|, i en ambdós casos s’han obtingut e-values significatius (de l’ordre de e-09 i e-10).
Per aquesta família no es coneix un motiu concret conservat, però tant en SelS com en SelK, la selenocisteïna es troba en les últimes posicions de la seqüència: en la SelS és la penúltima i en la SelK és l’avantpenúltima. Les hem trobat perfectament alineades tant amb la query humana com amb la d’elefant.
Quan s’ha fet la cerca d’elements SECIs se n’han identificat en aquesta regió.
Pel que fa als homòlegs en cisteïna, la seqüència de SelS trobada al SelenoDB és més curta que la de la selenoproteïna i no conté el fragment on trobaríem la selenocisteïna (en aquest cas hauria de ser una cisteïna). Degut a aquest fet, no podem afirmar que tinguem un homòleg en cisteïna de la SelS. Cal destacar que trobem elements SECIS en aquesta regió, però això és degut a que es troba en el mateix scaffold que la selenoproteïna.
SelP
Aquesta selenoproteïna és la més abundant en plasma, després de GPx3, i s'estima que conté el 50% del seleni present en el plasma sanguini. És secretada al torrent sanguini pel fetge en forma de glucoproteïna i s'encarrega de transportar el seleni als diferents teixits del cos. També s'hipotetitza que pot servir com a quelant de metalls pesats i que pot tenir activitat antioxidant.[17]
És l'única selenoproteïna eucariota que presenta múltiples selenocisteïnes a la seva seqüència, per la qual cosa és un bon reservori de seleni, i per tant és capaç de transportar-lo per tot l'organisme. Presenta un residu de selenocisteïna a l'extrem N-terminal, l'extrem C-terminal presenta múltiples selenocisteïnes, i a la part central hi trobem regions riques en histidina.
En aquest cas, hem aconseguit alinear bastant bé la SelP, tant ulitzant la query d’humans com la query de l'elefant. En ambdós casos ens falta un tros del final, però la resta s'alinea molt bé, amb un score de 99, i un grau d'homologia del 81,85% amb la query de H. Sapiens i del 89,71% amb la query de L.africana. Es troba en el scaffold gi|406062180|gb|JH864143.1| del genoma de O. afer i en ambdós casos el e-value és significatiu, essent de l'ordre de e-11 i e-19, respectivament.
S'han identificat múltiples selenocisteïnes en la seqüència proteica de O. afer, conservant-se l'estructura explicada anteriorment, amb un residu en l'extrem N-t, seguit d'una regió rica en histidines, i una regió C-t rica en selenocisteïnes, algunes de les quals s'alineen amb cisteïnes en la query corresponent. En el cas de L. africana hi ha més alineaments coincidents entre selenocisteïnes respecte els humans, on trobem més alineaments de selenocisteïna-cisteïna. Això s'explica perquè L.africana és més proper evolutivament a O. afer que H. sapiens, i per tant han divergit menys.
SelM
SelM és una selenoproteïna íntimament relacionada amb Sel15, ja que com aquesta, conté un residu Sec la regió catalítica en la posició N-terminal, amb el motif -CXXU-. La funció d'aquesta proteïna encara no és clara, tot i que se sap que desenvolupa un paper molt important al cervell. S’ha vist que en cas de sobreexpressió de la proteïna Presenilina, relacionada amb l’aparició prematura d’alzheimer, SelM queda quasi totalment inhibida. A part d’expressar-se al cervell, també ho fa en menor grau a cor, pulmó, ronyó, úter i placenta.[12]
En el nostre cas, els resultats mostren que trobem SelM en el genoma de O. afer amb el característic motiu -CXXU- juntament amb una regió SECI, la qual ha estat descrita en la bibliografia com una variant rara amb citosina-citosina en el loop apical de SECIs, i que el nostre output del Seblastian confirma. L'scaffold de la proteina és gi|406062185|gb|JH864138.1| amb una homologia de 88,46 i un e-value de 6e-26 per a humans, i per a Loxodonta, 94,12% d’homologia i 5e-25 d’e value, que ens reafirma l’obtenció de resultats molt significatius.
FAMILIA MsrA i SelR
La família de proteïnes Msr (methionine sulfoxide reductase) són un grup de proteïnes que catalitzen la reducció de residus oxidats de metionina. Aquesta família és capaç de reparar els danys causat per l'oxidació gràcies a la seva activitat redox, i així, aconsegueix reparar enzims amb funció oxidativa que han estat danyats i ja no funcionen. La oxidació de l'aminoàcid metionina comporta la formació d'isòmers R i S, generant aquests dos tipus. S'ha demostrat que la família Msr protegeix els organismes contra processos d'estrès oxidatiu, entre els quals trobem la hipòxia i reperfusió, jugant un paper important en la determinació de la durada de vida cel·lular.
Hi trobem dues famílies, MsrA i MsrB, o SelR. La primera té una única isoforma en mamífers, la qual repara estereoespecíficament els isòmers S. La segona família, MsrB, conté tres isoformes: MsrB1 (SelR1) MsrB2 (SelR2) i MsrB3 (SelR3), que reparen estereoespecíficament els isòmers R. Ambdues famílies difereixen en seqüència genòmica i estructura proteica, i d'aquest fet s'infereix que han divergit evolutivament per arribar fer la mateixa funció. En SelR1 hi trobem en el domini catalític una Sec, i és per això que hem decidit estudiar tota la família de Msr, per així poder confirmar una homología de cisteïna entre O. afer i elefants/humans, o bé un nou canvi a Sec.
MsrA
Els nostres resultats mostren que MsrA s’alinea amb una homologia de 99,65% amb Loxodonta i amb humans la homologia és de 95,83%. S’ha vist que falta un fragment de la seqüència respecte humans, tant per davant com per darrere, mentre que amb Loxodonta s’alinea quasi a la perfecció. Això ens confirma que evolutivament, Loxodonta i O. afer són més propers que no pas amb humans. L’scaffold d’aquesta proteïna correspon a gi|406065373|gb|JH863771.1| i en tots dos casos l’e-value és significatiu, essent de 2e-22 per humans i per Loxodonta 9e-25. La seqüència mostra un alineament Cys-Cys de O. afer respecte elefants i humans en el centre catalític. Així, es confirma la homologia en cisteïna.
SelR1
Aquesta selenoproteïna conté una Selenocisteïna en el domini catalític. L’alineament presenta una homologia de 89,54 i 84,22% en humans i Loxodonta, respectivament, i un e-value de 1e-23, i 2e-12, i la trobem present en l’scaffold gi|406065284|gb|JH863779.1|. Tal i com mostren els resultats, doncs, en O.afer aquesta proteïna es manté, conservant la Sec en el seu domini catalític, juntament amb la regio SECIS predita per SeciSearch3.
SelR2
Aquesta proteïna es localitza exclusivament al mitocondri i la podem classificar com a homòleg en cisteïna. La literatura mostra que aquesta proteïna, i la isoforma SelR3, explicada a continuació, contenen en el seu domini catalític un motiu molt conservat (-CINS-) tant en humans com en Loxodonta. Els nostres resultats mostren que en O. afer també es conserva de forma total el motiu esmentat, amb la cisteïna corresponent. La proteïna es localitza en l’scaffold gi|406066056|gb|JH863753.1| i l’alineament mostra una homologia de 100% i 94,76 respecte Loxodonta i humans, mentre que l’e-value és de 3e-13 i 9e-20, respectivament. Predim un element SECI tot i no tractar-se de selenoproteïna. Això pot ser degut a la similaritat de seqüència entre aquesta proteïna amb d’altres de la mateixa família que sí que són selenoproteïnes.
SelR3
A diferència de l’anterior, aquesta proteïna es localitza al reticle endoplasmàtic, però conté el mateix motiu -CINS- amb la seqüència conservada totalment en O. afer. L’alineament amb la query de H.sapiens mostra una homologia del 100% i un e-value de 7e-21. Per a Loxodonta, trobem una homologia de 89,29% i un e-value de 7e-21. Es localitza a l’scaffold gi|406064754|gb|JH863815.1|, en la qual s’han predit tres SECIs, segurament degut a la homologia amb SelR1.
SelU
Aquesta selenoproteïna es va trobar inicialment en eucariotes unicel·lulars, peixos i ocells. En mamífers superiors, la SelU es troba en la forma amb cisteïna i no amb selenocisteïna. Hi ha tres subfamílies de SelU i en la única on es troba selenocisteïna és la SelU1. En el cas d’aquesta subfamília, s’ha vist que el canvi de selenocisteïna per cisteïna es va donar en l’estadi de mamífers primitius. S’ha observat que aquestes proteïnes formen part de la superfamília de proteïnes tioredoxines. [18]
En el nostre cas, hem trobat 3 homòlegs en cisteïna (SelU1, SelU2, SelU3) en l’anàlisi amb la query humana. En el cas de SelU1, aquesta es troba en l’scaffold gi|406064146|gb|JH863881.1|, amb una homologia del 100% i un e-value de 4e-57. En el cas de SelU2, es troba en l’scaffold gi|406066985|gb|JH863723.1|, amb una homologia del 92.86% i un e-value de 1e-18. Finalment, la SelU3 es troba en l’scaffold gi|406064396|gb|JH863854.1|, presenta una homologia del 89.86% i té un e-value de 3e-29, fet que indica molta significància.
En el cas de les querys de Loxodonta només trobem l’homòleg de cisteïna en SelU3.Trobem aquesta proteïna en l’scaffold gi|406064396|gb|JH863854.1|, amb una homologia del 98.55% i amb un e-value de 6e-33.
Tornar a dalt
SELENOPROTEÏNES DE T.nigroviridis: Fep15, SelL i SelU
A part de buscar les Selenoproteïnes d’humans i de loxodonta en l’organisme d’estudi, l’O.afer, també hem buscat aquestes tres Selenoproteïnes de T.nigroviridis, ja que no es troben anotades en el genoma ni d’humans ni de loxodonta. Després de realitzar tots els programes pertinents, no hem trobat cap d’aquestes tres selenoproteïnes en el genoma de l’Orycteropus afer.
En el cas de la Fep15, quan fem el blast no obtenim cap hit amb un e-value suficientment significatiu, és per això que hem conclòs que no tenim la proteïna en l’organisme estudiat.
Pel que fa a la SelL, no obtenim cap hit amb el blast, de manera que no s’alinea cap seqüència del genoma de l’O.afer amb aquesta selenoproteïna.
En el cas de la SelU, sí que hem obtingut hits significatius amb el blast, però la proteïna predita en el genoma de l’O.afer no conté cap selenocisteïna i, conseqüentment, no hi trobem cap element SECIS. El fet de que no hi hagi cap selenocisteïna podria indicar que aquest aminoàcid s’ha perdut evolutivament, ja que el T.nigroviridis és un organisme més antic que l’O. afer.
Tornar a dalt
MAQUINÀRIA TRANSCRIPCIONAL
Existeixen diferents proteïnes o estructures implicades en la síntesi de selenoproteïnes, aquesta són:
-
SBP2 (SECIS binding protein 2) →S’uneix a l’element SECIS i recluta el factor d’elongació eEFSec
-
eEFSec (SEC elongation factor) →Permet la interació del tRNA de la selenocisteïna amb SBP2 i aproxima el tRNA de transferència al codó UGA del mRNA de la selenoproteïna, per a que aquest pugui incorporar la selenocisteïna i seguir amb la traducció.
-
tRNAsec → és el tRNA encarregat d’incorporar la selenocisteïna en la cadena polipeptídica.
-
SPS1 i 2 (selenofosfat sintetases 1 i 2)→ sintetitzen selenofosfat a partir del seleni.
-
PSTK (fosfoseril tRNA quinasa) →fosforila el seril-tRNA a O-fosfoseril-tRNA, que és un intermediari actiu per a la síntesi de selenocisteïna.
-
SLA/LP o SecS (selenocisteïna sintasa)→converteix O-fosfoseril-tRNA en selenocistenil-tRNA, necessari per a la síntesi de selenoproteïnes.
-
SECp43 (proteïna associada a selenocisteïna 1)→ Té un paper important en els primers passos de la síntesi de selenocisteïna i en la càrrega del tRNA corresponent que permetrà la incorporació de la selenocisteïna a les selenoproteïnes. Estabilitza el complex SBP2 - eEFSec - tRNA afavorint l’eficiència de la síntesi de selenoproteïnes. [11]
Hem analitzat la presència de maquinària de síntesi de selenoproteïnes en el genoma d’Orycterpus afer utilitzant les querys de la maquinària d’humans i de Loxodonta. Hem aconseguit identificar SBP2, eEFSec, SPS 1 i 2, PSTK, SECp43 i SecS, és a dir trobem tota la maquinària de transcripció necessària per la síntesi de selenoproteïnes cosa que augmenta la probabilitat que les nostres prediccions siguin certes.
Tornar a dalt
|