
La forma biològica més rellevant del seleni és l’aminoàcid selenocisteïna (Sec) (1). De fet, el fenomen que defineix les selenoproteïnes és la inclusió d’aquest residu. L’aminoàcid 21 es diferencia del seu anàleg, la cisteïna (Cys), en la substitució d’un àtom de sulfur (Cys) per un àtom de seleni (Sec). El seleni té propietats químiques úniques (2) que doten a les selenoproteïnes d’una gran importància funcional i les fan imprescindibles per a l’organisme.
Tot i que moltes de les funcions de les selenoproteïnes descobertes fins al moment no es troben ben caracteritzades, se’ls atribueixen rols en processos RED-OX, en el metabolisme d’hormones, síntesi de fosfolípids, correcte plegament de proteïnes en el RER, entre altres (3–5).
A més, s’ha descrit les seleproteïnes com a possibles marcadors evolutius (3). A partir de les diferències al selenoproteoma de diferents espècies, podem establir un arbre filogenètic, en el qual s’il·lustren diferents esdeveniments evolutius. En aquest context, creiem rellevant la correcta anotació de les selenoproteïnes. Una major informació sobre el conjunt d’aquestes proteïnes serà d’utilitat per a futurs estudis relacionats.
L’espècie Propithecus coquereli, un primat endèmic de l’illa de Madagascar, ha estat recentment seqüenciada. Els lèmurs són un infraordre (Lemuriformes) de primats que va sorgir en aquesta illa en particular, patint un procés evolutiu de coll d’ampolla i, a continuació, una radiació adaptativa (6), molt diferent de primats continentals (6). Per tant, considerem Prophitecus coquereli com un bon model per a l’estudi d’una població amb un patró de diversificació genètica propi de les espècies endèmiques de les illes.
El nostre projecte s’ha centrat en l’anotació de les selenoproteïnes de Prophitecus coquereli. Per obtenir els nostres resultats hem alineat el genoma del lèmur amb les selenoproteïnes d'Homo sapiens sapiens. L’humà és un primat i, per tant, una espècie propera al nostre subjecte d’estudi. Així mateix, dins del grup dels primats, el seu genoma és el més ben estudiat i, en conseqüència, el més ben anotat.
El selenoproteoma humà està format per 25 selenoproteïnes (7). D’acord amb els següents criteris que defineixen una selenoproteïna (codó UGA + presència d'element SECIS), hem trobat 25 selenoproteïnes al genoma de Prophitecus coquereli. Els nostres resultats coincideixen amb el contingut de selenoproteïnes esperat per a mamífers placentaris, concretament primats (3).
Família DI
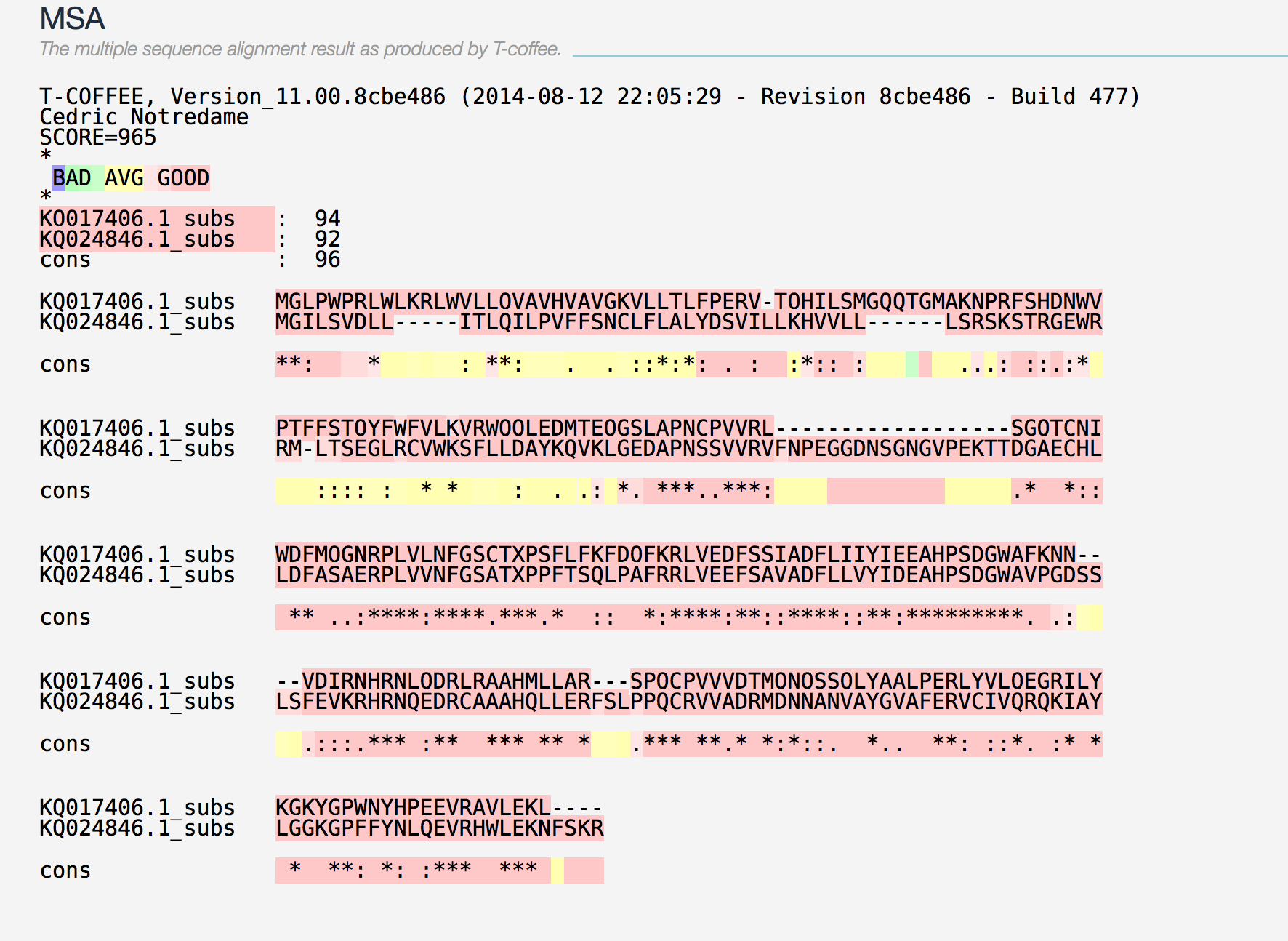
La famíla de les Iodotironina deoidinases (DI) presenta una alta homologia intrafamiliar (3). Trobem homologia en les mateixes regions del genoma de P.coquereli per a les proteïnes DI1, DI2 i DI3 d’humà (Taula.1). També es pot afirmar que es tracta d’una família molt conservada entre les dues espècies, de fet, trobem més homologia entre les proteïnes d’humà i les de P.coquereli (SCORES 999-1000) que entre les de P.coquereli contra elles mateixes (SCORES 936-965) i les d’humà contra elles mateixes (SCORES 931-966). Podeu veure els resultats a: DI1-DI2 , DI1-DI3 , DI1H-DI3H , DI1H-DI2H .
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Hem comprovat que la proteïna contra ella mateixa dóna un SCORE de 1000. Podem dir que es dóna una major homologia interespecífica que intraespecífica. En principi, aquest fet contradiu l'homologia intrafamiliar que s’ha atribuït a les DI, tant per a P.coquereli com per a H.sapiens. Tot i així, els valors de SCORE dels alineaments no mostren diferències estadísticament significatives (p-values entre 0.65 i 0.66).
El fet que aquesta família estigui molt conservada pot ser degut a que les tres proteïnes tenen funcions de regulació del metabolisme tiroideu, passen de la prohormona T4 a la hormona acitva T3, mentre que la DI3 la inactiva (8–11), participant en processos de metamorfosi i termogènesi, indicant que es troben presents des de la separació d’amfibis, com a mínim.
La regió més conservada és la de la selenocisteïna (9), de fet, podem veure que en les imatges dels alineaments anteriors com, efectivament, hi ha una major conservació al voltant de la selenocisteïna. Això indica que la Sec té un paper principal a la funció de la proteïna.
Els resultats de l’alineament mitjançant el BLAST per a DI2 i DI3 mostren un hit per a cada una d’aquestes proteïnes amb un E-value < 10-100 (10-13 i 10-176, respectivament). Deduïm que aquest valor estadístic ve donat per la longitud del hit (202 i 265 aminoàcids, respectivament). Aquesta es correspon amb la longitud d’un dels exons. (Taula.2)
DI2 té un segon open Reading frame (codó AUG) que es pot observar a la proteïna de P.coquereli. També s’observa que, a més de la Sec conservada entre Homo sapiens i P.coquereli, trobem a la proteïna d’humà una segona Sec localitzada a prop de l’extrem 3’UTR (4) trobant a P.coquereli un gap. No pot ser degut a una pèrdua a l’extrem 3’UTR ja que hi trobem dos elements SECIS al 3’ de la proteïna. Podria ser que s'hagués donat el canvi d’una Sec a un STOP, a l’alineament veiem que el primer aminoàcid no codificat a l’extrem 3’ és el que a la proteïna d’humà trobem una Sec. Podem veure els tres fenòmens a l’alineament per T-COFFEE .
DI3 ha estat descrita com a proteïna intronless (3), de fet, així la trobem al nostre organisme subjecte tant pel que fa al resultat del BLAST com al resultat de l’EXONERATE. (Taula.1) (Taula.2)
Família GPx
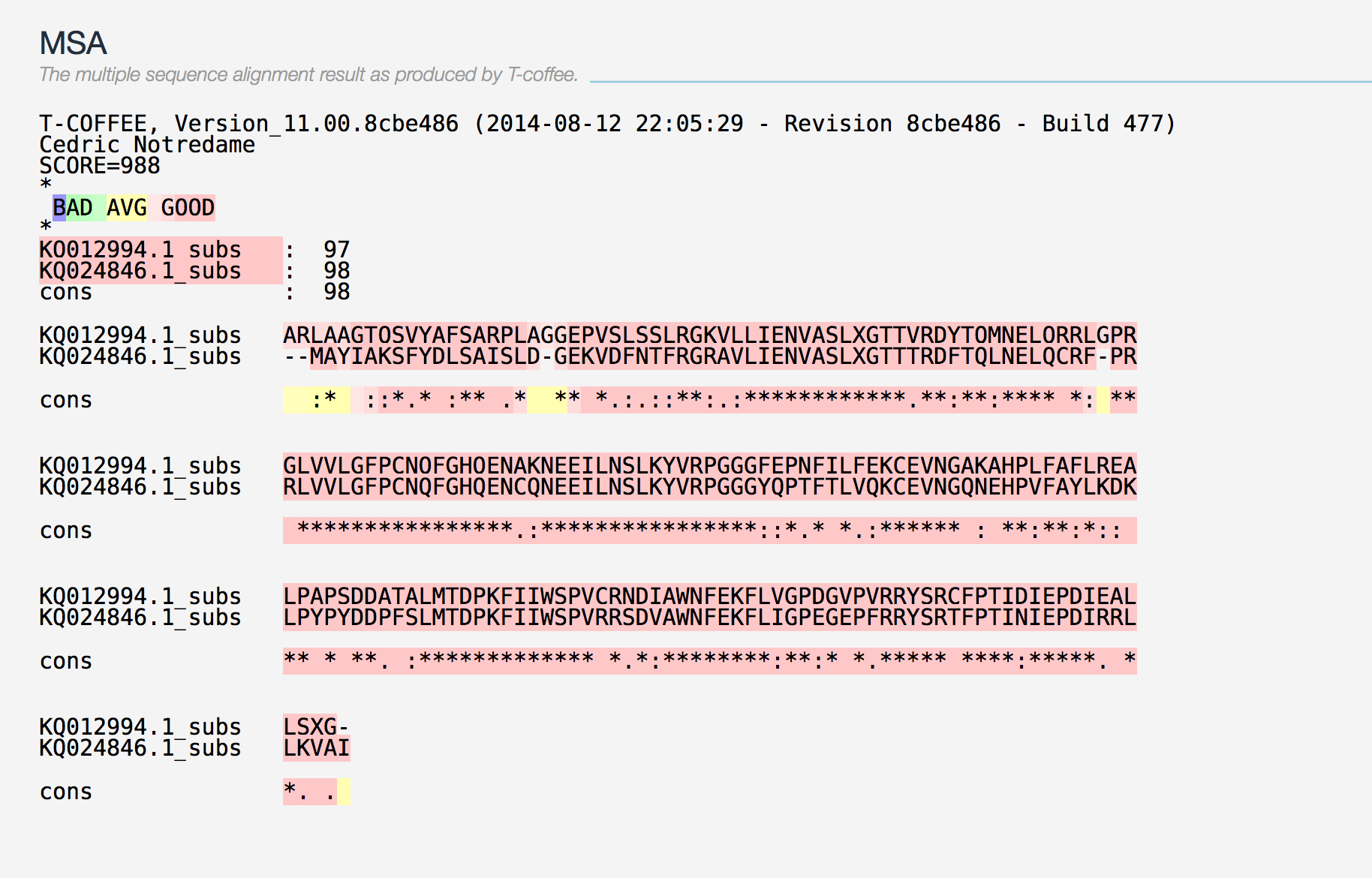
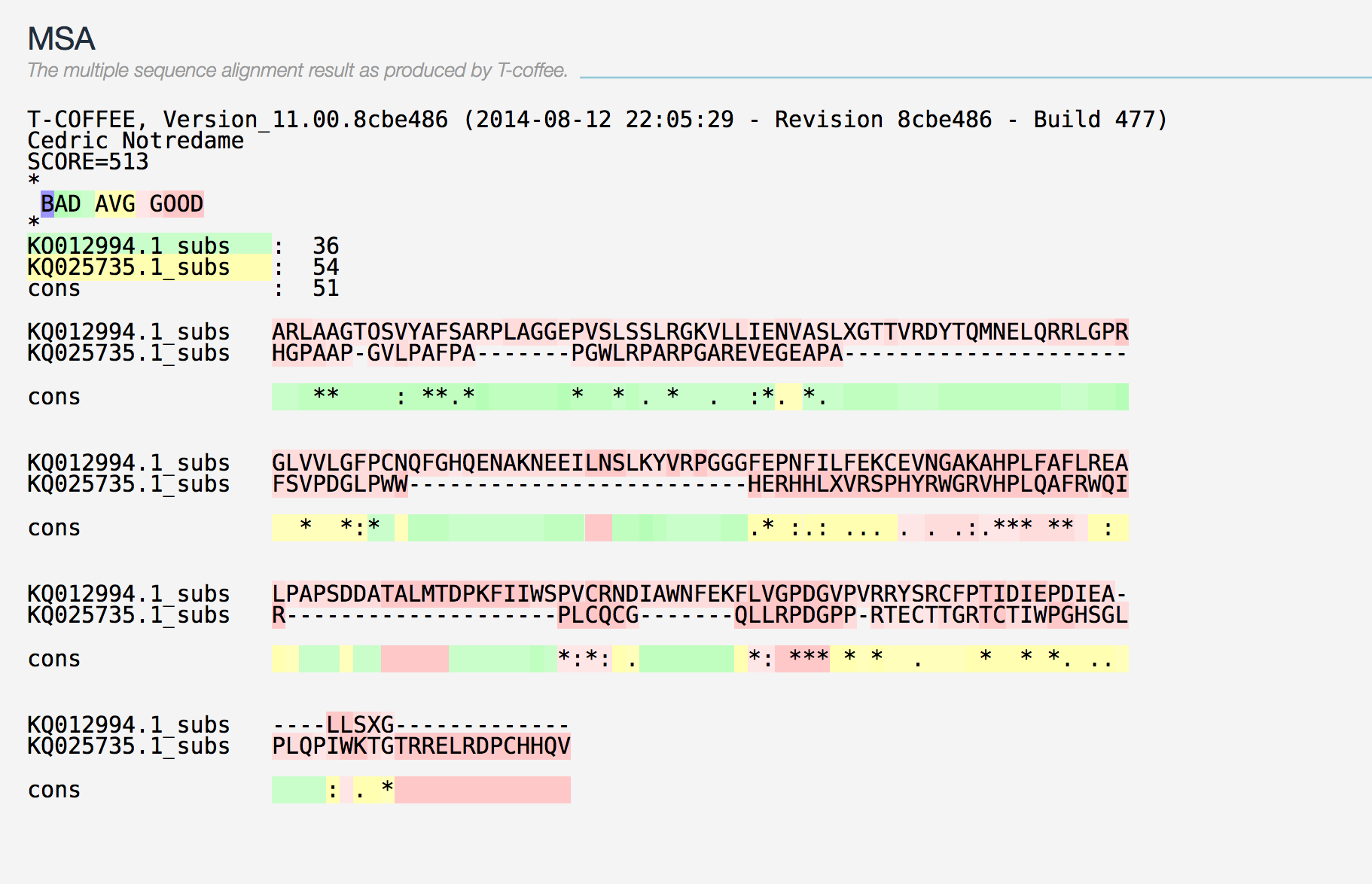
En mamífers trobem 8 proteïnes glutatió peroxidasa homòlegs molt conservades, dels quals 5 són selenoproteïnes (GPx1, GPx2, GPx3, GPx4, GPx6 en humà). Aquest grup d’enzims amb activitat peroxidasa té com a principal funció biològica protegir els organismes del dany oxidatiu (12–15). En referència a la filogènia, la família de les GPx consisteix en tres grups evolutius que descendeixen d’un ancestre comú amb Cys: GPx1/GPx2, GPx3/GPx5/GPx6 i GPx4/GPx7/GPx8 (3, 15–17). GPx5 i GPx6 van aparèixer en mamífers placentaris com a resultat d’una duplicació en tàndem de GPx3 (3, 15). Hem comparat dos alineaments realitzats amb TCOFFEE per a les proteïnes GPx1-GPx2 i GPx1-GPx3, observant diferències a l’homologia d’ambdós alineaments (SCORES: 988 i 513 p-value: 7.716038e-21), dades que recolzen les evidències prèvies referents a l’agrupament filogenètic d’aquesta família.
{kind=link}
{kind=link}
Gran part de les proteïnes d’aquesta família posseeixen pseudogens al genoma d'H.sapiens (GPx1:3, GPx2:1, GPx4: 4) (3). Consideraríem que esdeveniments genòmics d’aquest tipus donarien hits a l’output del BLAST que cobrissin un alt percentatge de la llargada de la proteïna, i presentessin uns valors estadístics (d’identitat i E.value) significatius, tot i així, als nostres resultats no hi trobem cap cas d’aquestes característiques. No podem confirmar la presència de pseudogens al genoma de Prophitecus coquereli.
GPx1 a més de tenir una Sec a la mateixa posició que a la proteïna humana, a la target hi trobem una Sec més a l’extrem 3’ que a la query la qual és un residu de Q. En canvi, a GPx2 hi trobem una Sec que coincideix en la posició de la target i la query. Deduïm que la segona Sec trobada a la seqüència de GPx1 ha de tenir alguna funció degut a que la seva síntesi suposa una despesa energètica extra per a la cèl·lula.
GPx4 és una selenoproteïna de 7 exons tant en humà (15) com en P.coquereli, com indiquen els nostres resultats. A l’humà existeixen tres isoformes, que no trobem de manera clara en el lèmur. La selenocisteïna es troba a la mateixa posició en les dues espècies.
De moment GPx6 no ha estat purificada, i no existeixen estudis cinètics, per tant el coneixement d’aquesta GPx és encara limitat (15). Ha sigut reportat un element SECIS “fòssil” (15), aquest no el trobem al genoma del nostre animal d’estudi. Trobem una Sec, però l'absència de l’element SECIS podria implicar la no funcionalitat d’aquesta, no sent GPx6 una selenoproteïna, fet que ha estat observat en altres espècies de mamífers.(3)
Família Sel:
SelM, Sel15 i SelH: es tracta de tres proteïnes del reticle endoplasmàtic considerades homòlogues llunyanes pel fet que comparteixen un 31% d’identitat de la seqüència. Presenten dos motius, -Cys-X-X-Sec- i -Cys-X-Sec, en humà, que també es pot observar a Prophitecus coquereli (alineaments: SelM Sel15 SelH), i un plegament típic de proteïnes com les tioredoxines.
SelM: s’expressa principalment al cervell on s’ha relacionat amb efectes neuroprotectors. A més, s’ha proposat com a possible enzim antioxidant per la seva implicació en la regulació de l’alliberament de Ca2+ del reticle endoplasmàtic en resposta a H2O2. Tot i així, la seva funció exacta i el rol que desenvolupa en la fisiologia del cervell encara s’ha d’acabar d’establir.
Sel15: s’ha vist associada amb UDP-glucosa, una glicoproteïna glucosiltransferasa del reticle endoplasmàtic amb la funció de modular el correcte plegament de les proteïnes.
Sel H: selenoproteïna similar a les tioredoxines que conté un motiu conservat -Cys-X-X-Sec- (els nivells de mRNA de Sel H són molt sensibles a la ingesta de Se) (4). Sel H es troba molt expressada en etapes relacionades amb el desenvolupament de l'embrió i després en una gran varietat de teixits. Es localitza al nucli, on alguns estudis suggereixen que du a terme una activitat redox unida al DNA, regulant els nivells d’expressió de gens involucrats en estats d’estrès oxidatiu.
Sel I: s’ha determinat la seva relació amb les vies de biosíntesi dels fosfolípids de la majoria de teixits. Concretament, Sel I posseeix un domini CDP-alcohol fosfatidil transferasa conservat que és comú en les colina fosfotransferases (CHPT1) i les etanolamina fosfotransferases (CEPT1) (4). La seqüència d’aminoàcids de Sel I conté 7 hèlix transmembrana.
Nosaltres hem trobat tres hits amb valors d’identitat i E-value significatius, i que per tant hem considerat adequat analitzar-los independentment. El primer d’ells, que suposem que es tracta del hit de la proteïna sencera, posseeix múltiples exons i coincideix en la posició de la Sec amb la query d'H.sapiens.
El segon conjunt de hits analitzat dóna una senyal de 100 aminoàcids amb un E-value de 10-17 i un altre de 300 aminoàcids aproximadament amb un E-value de 10-171, formant una proteïna de dos exons. El tercer conjunt de hits presenta també valors molt bons. Totes aquestes dades ens impulsen a pensar que podria tractar-se de l’existència descrita prèviament d’homòlegs molt similars al genoma de P. coqueleri (3), fet que no coincideix amb el patró de diversificació radial al que es va sotmetre aquesta espècie. També podria estar degut al fenomen de retrotransposició (18–20). Aquest esdeveniment dóna lloc a repeticions al genoma de parts del gen duplicat, que es poden produir en uns hits com aquests. Tots aquests hits els considerem possibles selenoproteïnes.
Sel K: proteïna transmembrana que es troba al reticle endoplasmàtic. A nivell de les cèl·lules del sistema immunitari, és important per al flux d’ions Ca2+, per a la proliferació de cèl·lules T i la seva migració juntament amb la dels neutròfils. A més, es troba involucrada en el procés de degradació de proteïnes glicosilades associades al reticle endoplasmàtic (ERAD), és a dir, participa en l’eliminació de proteïnes del citoplasma que no s’han plegat correctament al reticle endoplasmàtic. La seva expressió és elevada en teixits com: el cor (sobreexpressada protegeix als cardiomiòcits de l’estrès oxidatiu) i a la melsa. Es tracta d’una proteïna petita (94aa) que està considerada molt antiga dins de les selenoproteïnes (3). En P.coquereli trobem a la mateixa regió del genoma dos hits molt bons, la qual cosa pot significar que hi ha un domini molt conservat d'aquesta (possiblement la seva regió transmembrana).
Sel N: proteïna transmembrana similar a Sel K i Sel S que es troba a la membrana del reticle endoplasmàtic i que presenta dues isoformes. Mentre la isoforma 1 es correspon amb el transcrit sencer, la isoforma 2 està formada per 3 exons menys, eliminats al procés d’splicing en humà, per tant no podríem detectar aquesta segona isoforma al nostre estudi, de fet trobem només un conjunt de hits localitzats a la mateixa regió genòmica.
Totes dues isoformes han estat detectades al múscul esquelètic, al cervell, als pulmons i a la placenta on predomina el transcrit de la isoforma 2. S’ha trobat un link entre mutacions en el gen de la Sel N (SEPN1) i l’aparició de desordres musculars coneguts amb el nom de miopaties de tipus SEPN1 (4). A P.coquereli ens donen molts hits, significant que té moltes seqüències repetides d'aquesta proteïna en el genoma.
Sel O: Malgrat haver estat descoberta fa més d’una dècada és de les selenoproteïnes menys caracteritzades. Es considera una proteïna distribuida extensament perquè té homòlegs en animals, bactèries, llevats i plantes. Tot i així, la seva funció als organismes segueix sent una incògnita. Només s’ha detectat selenocisteïnes als homòlegs de vertebrat, localitzades en la penúltima posició de l'extrem C-terminal (3). En homòlegs d’altres organismes la selenocisteïna està reemplaçada per una cisteïna, en canvi, en P.coquereli sí que la trobem. (Alineament Sel O )
Sel P: es tracta d’un membre únic dins la família de les selenoproteïnes ja que conté múltiples residus de selenocisteïna en una sola molècula. Concretament, s’ha pogut identificar fins a 10 residus Sec en la Sel P d’humans i ratolins. En P.coquereli, al no poder córrer l'EXONERATE en aquesta proteïna, ens vam basar en els resultats del BLASTOUT. En aquest veiem, per un hit, la presència de 7 Sec. Al ser un Blastout no podem saber del cert si n'hi ha exactament 7. A més, d’un 40% a un 50% del Se en plasma es troba en la selenoproteïna P (4). Per aquest motiu, se li atribueix la funció de transport del Se obtingut per la dieta. De tal manera que és el responsable de regular l'homeòstasi del seleni a l'organisme, es sintetitza al fetge i porta Se a determinats òrgans i teixits. Recentment, s’ha associat l'Alzheimer a partir de l'anàlisi de teixit post-mortem on s’ha trobat que Sel P i les plaques amiloides colocalitzen.
Sel R: forma part de la família dels pèptids metionin sulfòxid reductases, sent l'única de les proteïnes que conté seleni. Sel R també anomenada MsrB, no comparteix homologia ni en estructura ni en seqüència amb MSrA, un altre membre de la família. En conseqüència, es considera que tenen orígens evolutius diferents. Aquests pèptids reparen proteïnes que han estat inactivades per l’oxidació de metionines, catalitzen la reducció de les metionines-sulfòxid a metionines. Sel R té diverses isoformes: Sel R1, Sel R2, Sel R3 (Sel R2 i Sel R3 presenten més homologia). Al genoma de l’espècie estudiada trobem un sol hit que podria considerar-se significatiu com per relacionar-ho amb una altra isoforma.
Sel S: és una proteïna transmembrana del reticle endoplasmàtic expressada en múltiples teixits. S’ha proposat que podria participar en l’eliminació de proteïnes mal plegades del lumen del RE i de protegir les cèl·lules del dany oxidatiu i l’apoptosi induïda per l’estrès del RE (4). S’ha demostrat com alguns SNPs en el promotor de Sel S regulen els nivells de citoquines proinflamatòries. Malalties cardiovasculars, atacs de cor, preclàmpsia, artritis reumatoide així com el càncer d’estómac, són algunes de les malalties relacionades amb variacions genètiques de Sel S. Igual que Sel K, Sel S presenta una Sec en la seqüència C-terminal i una sola regió transmembrana a l’extrem N-terminal en humà. Contràriament, no hem trobat cap Sec a la proteïna de P.coquereli, en cas que aquest residu fos necessari per la correcta funció de la proteïna trobaríem que aquesta funció estaria seleccionada negativament en aquesta espècie. Considerem, per tant, que Sel S no és una selenoproteïna.
Sel T: aquesta proteïna forma part d’una família de selenoproteïnes amb característiques redox (Sel W, Sel H i Sel V). Tots els membres tenen plegament de tipus thioredoxin-like (TRX) i un motiu molt conservat -Cys-x-x-Sec- (4). En base a aquests trets s’ha predit que el seu rol és semblant a les proteïnes oxidoreductases però la funció exacta que desenvolupen es desconeix actualment. La Sel T s’expressa tant en el desenvolupament embrionari com en l’edat adulta i es localitza al reticle endoplasmàtic mitjançant un domini hidrofòbic. A P.coquereli, a més de la Sec que està conservada amb la query, hem trobat una segona Sec més a prop de l’extrem 5’.
Sel V: és una de les selenoproteïnes menys caracteritzada. Recentment, es creu que ha evolucionat per la duplicació de Sel W i es troba només en mamífers placentaris. Tot i així, s’ha pogut estudiar la seva pèrdua en alguns organismes concrets com els goril·les (3). Sel V està estretament relacionada amb Sel W però té una mida més gran degut a un domini NH2-terminal del qual no se’n coneix la funció, a P.coquereli trobem que el hit òptim de Sel V es localitza en un dels hits poc significatius de Sel W, confirmant que Sel V prové de Sel W. La seva expressió es dóna només als testicles, per la qual cosa s’ha hipotetitzat que podria estar involucrat en la reproducció.
Sel W: és una de les selenoproteïnes més petita i abundant en mamífers. Va ser una de les primeres proteïnes identificades per contenir una selenocisteïna. Es localitza al citosol i té alts nivells d’expressió als músculs i al cervell. En ser purificada del múscul de ratolí es va poder observar com formava part d’un complex amb glutatió. S’ha proposat que Sel W podria estar involucrada en la regulació d’estats redox. Malgrat tot, el seu rol en la fisiologia dels organismes encara no s’ha determinat. Com hem dit anteriorment, hem trobat que Sel W té un hit a les coordenades del hit òptim de Sel V. Tot i així el e-value d’aquest hit no és significatiu. Degut a que aquesta proteïna tenia diversos hits molt bons hem considerat adient comptar-los tots ells com a selenoproteïnes.
Família SPS2
SPS2: És un enzim involucrat en la síntesi de selenoproteïnes. De fet, autoregula la seva producció junt amb la de la resta de selenoproteïnes. Catalitza la síntesi del selenofosfat, un donador actiu de seleni que es precisa per la síntesi de selenocisteïnes. En referència als 4 hits que hem trobat en aquesta proteïna al genoma de P.coquereli, podrien ser deguts a una duplicació que s’hagi mantingut, augmentant la complexitat de la maquinària. Degut a que aquesta proteïna tenia diversos hits molt bons hem considerat adient comptar-los tots ells com a selenoproteïnes.