|
 |
| |
|
| |
|
| |
El seleni és una substància natural i sòlida que es troba molt present a l’escorça terrestre tot i que de manera irregular. Els seus nivells són molt baixos dins l’organisme, però és incorporat a les proteïnes a través d’un sistema genètic específic. El Seleni és indispensable per als éssers vius, i el seu dèficit pot ser la causa de determinades malalties. Tanmateix, la seva presència en excés pot tenir efectes tòxics.
|
|
| |
Puja dalt |
|
| |
|
|
| |
|
|
| |
|
| |
Les selenoproteïnes són proteïnes la seqüència de les quals conté una selenocisteïna (Sec o U). Es troben en tots els organismes animals però no en les plantes (aquestes presenten una versió anomenada homòlegs de cisteïna, que són capaces de realitzar la mateixa funció que les Sec). Les selenocisteïnes es diferencien de les cisteïnes perquè presenten un àtom de seleni en comptes del radical sulfur, i són considerades el 21è aminoàcid.
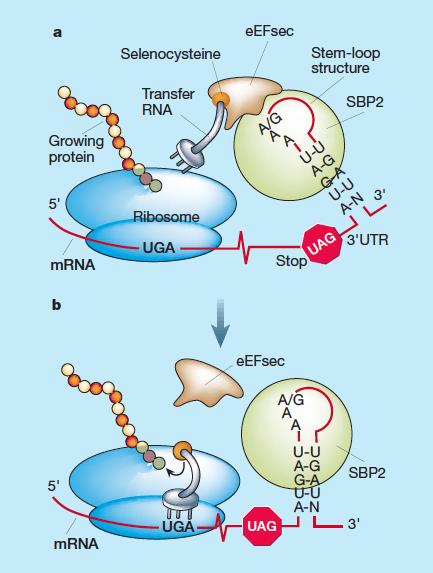
Són traduïdes pel codó UGA, que és suposadament un codó STOP i per tant normalment hauria d’aturar el procés de traducció. Però en el cas de les selenoproteïnes, una estructura tridimensional anomenada SECIS (Seleno Cystein Insertion Sequence), situada a la regió 3’ UTR del seu mRNA, possibilita que enlloc d’aturar la síntesi de la proteïna, el ribosoma afegeixi una Sec i continuï traduint fins a trobar qualsevol altre codó STOP. Així doncs, el codó UGA es recodifica per tal que es pugui introduir l’aminoàcid selenocisteïna al pèptid, fet que és possible gràcies a la maquinària especial de traducció de les selenoproteïnes.
La seqüència SECIS, que presenta una estructura secundària de bucle, permet la unió de SBP2 (Secis Binding Protein 2). Aquesta, al seu torn, s’encarrega de reclutar el tRNA específic de la selenoproteïna, i el procés de traducció continua fins al següent codó STOP. Finalment, es necessita el factor d’elongació eEFSec (Eukaryotic Elongation Factor Selenocysteine–specific), que interacciona específicament amb el Sec–tRNASec a través de SBP2, amb el qual forma un complex. Així doncs, SBP2 funciona com a pont entre el tRNA i el factor d’elongació. La seqüèncoa SECIS no es tradueix en la síntesi de la selenoproteïna.
En la majoria dels casos les selenoproteïnes actuen com a proteïna redox (defensa antioxidant, senyalització redox, oxidoreduccions...) tot i que també se’ls atribueixen funcions biològiques importants com intervenir en el metabolisme d’hormones tiroidees, implicacions en la resposta immunitària... Actualment hi ha 25 selenoproteïnes descrites en humans.
|
|
|
|
|
|
Síntesi d’aminoàcidsa
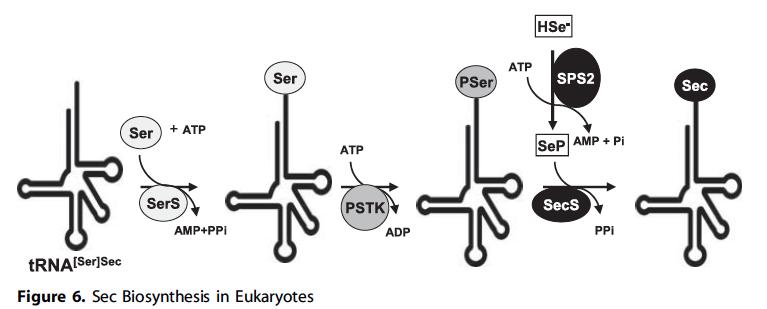
La selenocisteïna es sintetitza directament en el propi tRNA a partir d’una serina que actua com a intermediari. L’aminoàcid no es troba lliure a la cèl·lula ja que sinó podria ser incorporat erròniament en altres proteïnes. La serina interacciona amb aquest tRNA, anomenat Sec–tRNA, gràcies a la tRNA Seryl–tRNA sintetasa, donant lloc així al Seryl–tRNA (Ser–tRNASec). La serina serà el precursor de la selenocisteïna. Aquest complex és fosforilat per la PSTK (Phosphoseryl–tRNA kinase), i a continuació aquest fosfat és substituït per un donador de seleni, anomenat H2Se–P, per acció de la selenocisteïna sintetasa (SecS). Aquest donador de seleni, al seu torn, ha estat sintetitzat per la selenofosfat sintetasa 2 (SPS2). Per tant, la selenocisteïna sintasa, que consta d’un complex proteic format per les proteïnes Secp43 i SPS1, permet la incorporació del selenofosfat a l’aminoàcid.
|
| |
Puja dalt |
|
| |
| Famílies de Selenoproteïnes |
|
|
|
Conceptea
Actualment, encara hi ha moltes proteïnes pendents d’anotar en la majoria de genomes. El problema rau en que molts programes interpreten el codó UGA com a codó STOP i per tant ometen l’existència d’aquestes proteïnes. En conseqüència, per tal de trobar–les, cal utilitzar programes específics que detectin un perfil d’exó determinat amb el codó UGA i un element SECI situat a la regió 3’ UTR de l’mRNA. Una estratègia alternativa és fer el reconeixement mitjançant l’alineament de seqüències, ja que teòricament les selenoproteïnes estan ben conservades al llarg de l’evolució i presenten homologia entre diferents espècies. Aquesta segona alternativa és la que hem seguit al nostre estudi.
En el llistat següent es descriuen les característiques bàsiques de les selenoproteïnes que consten en el genoma d’humans i de la majoria de mamífers:
|
| |
Selenoproteïna Sep 15a
La selenoproteïna 15 és una proteïna tioredoxina–like localitzada al reticle endoplasmàtic. En el seu centre actiu presenta el motiu – Cys–X–X–Sec – (CXXU), el qual es troba altament conservat en determinades proteïnes. Malgrat que la seva funció no està del tot clara, determinats estudis en ratolins han demostrat com aquesta podria estar implicada en processos redox així com en el control del plegament de proteïnes. A més, es tracta d’un locus que de forma comú es troba mutat en càncers humans, indicant així una possible relació amb processos cancerígens. La seva expressió es regula a través del seleni que incorporem amb la dieta així com per estrès al RE, i aquesta és majoritària en: fetge, ronyons, pròstata i glàndula tiroides.
|
| |
Selenoproteïna H (Sel H)a
La selenoproteïna H és una proteïna de la família de les Rdx 12 (Sel H, Sel, T, Sel V i Sel W) localitzada al nucli i que presenta el domini tioredoxina–like, amb un motiu altament conservat – Cys–X–X–Sec – (CXXU) que li confereix la seva activitat redox.
|
| |
Selenoproteïna I (Sel I)a
La selenoproteïna I presenta un domini molt característic, el qual s’anomena CDP–alcohol fosfatidiltransferasa i que és propi d’enzims colina fosfotransferases i colina/etanolamina fosfotransferases. Es tracta d’una proteïna que està poc estudiada però que degut als dominis que presenta podria estar implicada en la síntesi de fosfolípids.
|
| |
Selenoproteïna K (Sel K)a
La selenoproteïna K és una proteïna localitzada al reticle endoplasmàtic. La seva funció no està massa clara. Tot i així, determinats estudis semblen demostrar com podria estar implicada en processos redox, en la regulació del calci així com en la via de degradació de proteïnes.
|
| |
Selenoproteïna M (Sel M)a
És una proteïna que es localitza al reticle endoplasmàtic de les cèl·lules i s’expressa en diversos teixits, especialment a l’encèfal. Ha estat relacionada amb la prevenció de l’estrès redox i la regulació del calci.
|
| |
Selenoproteïna N (Sel N)a
Aquesta selenoproteïna, amb funció encara poc caracteritzada, és expressada al múscul esquelètic, cervell i pulmó entre d’altres. És capaç d’interaccionar amb el calci gràcies a dominis específics i es relaciona amb el correcte funcionament dels músculs. Sembla estar força expressada en l’etapa pre–natal.
|
| |
Família de les Metionina-R-Sulfoxid Reductasa (Sel R)a
És un enzim que catalitza la reacció en que el sulfòxid de metionina es transforma a metionina. Aquest enzim pertany a la família de les oxidoreductases, i el gen que la codifica es localitza al nucli i al citosol de la cèl·lula i s’expressa en diversos teixits de l’organisme. La família de les methionine–R–sulfoxide reductases està formada per tres membres (SelR1, SelR2 i SelR3) però només la primera és una selenoproteïna.
|
| |
Selenoproteïna O (Sel O)a
La selenoproteïna O és una proteïna que de forma general es creu que juga un paper important en la senyalització cel·lular. Presenta diferents funcions, d’entre les quals destaquem la de quinasa; ja que presenta activitat fosfotransferasa, així com també intervé en processos redox. Es troba tant en eucariotes com en bactèries.
|
| |
Selenoproteïna P (Sel P)a
Aquesta selenoproteïna és una glicoproteïna extracel·lular que conté la majoria de seleni del plasma. Es creu que la seva funció és transportar el seleni des del fetge fins a tots els teixits perifèrics, actua com a antioxidant i té dominis d’unió a l’heparina.
|
| |
Selenoproteïna S (Sel S)a
És una proteïna de membrana del reticle endoplasmàtic que contribueix a tornar les proteïnes mal plegades al citosol perquè siguin degradades. Està relacionada amb el control de la resposta inflamatòria donat que es creu que pot regular la producció de citoquines.
|
| |
Selenoproteïna T (Sel T)a
Es tracta d’una proteïna localitzada al reticle endoplasmàtic i aparell de Golgi. Està potencialment implicada en processos d’adhesió cel·lular i en la millora de gens oxireductors. La seva funció no està del tot definida però es sap que està més expressada en l’embriogènesi que en l’etapa adulta.
|
| |
Selenoproteïna V (Sel V)a
Aquesta selenoproteïna va originar–se per una duplicació del gen de la SelW. Pertany a la família Rdx12 juntament amb les selenoproteïnes SelH, SelT i SelW. La seva funció no està definida, es creu que s’expressa únicament en els testicles dels mamífers.
|
| |
Família de les selenoproteïnes W (Sel W)a
Protegeixen a les neurones de l’estrès oxidatiu durant el seu procés de desenvolupament. També són expressades al cor, i múscul esquelètic.
|
| |
Família Glutatiò Peroxidases (GPx)a
Les glutatió peroxidases són un conjunt d’enzims que tenen com a funció principal protegir l’organisme de l’efecte degradant dels compostos hidroperòxids formats de manera endògena.
De manera que es produeix una reacció d’oxidació del glutatió al glutatió disulfur amb el consum d’una molècula de peròxid d’hidrogen, el qual es redueix a aigua. Així, protegeixen les cèl.lules del dany oxidatiu.
Aquesta família de selenoproteïnes és la més àmpliament descrita en vertebrats. Presenten un elevat grau d’homologia entre elles, fet que ens fa pensar que provenen d’un gen ancestral el qual va duplicar–se al llarg de l’evolució.
En mamífers hi ha descrites fins a 8 GPx, de les qual GPx1, GPx2, GPx3, GPx4 i GPx6 són selenoproteïnes, aquesta última únicament es troba en certes espècies. La resta són proteïnes homòlogues amb cisteïna.
|
| |
Família Iodotironin Deiodinases (DI)a
Les iodotiroidines deionidadses són una subfamilia de les diodinades importants en la regulació de les hormones tiroidees. Concretament passen la T3 a la seva forma activa T4. Aquesta hormona és doncs essencial per al correcte metabolisme de l’individu. Trobem tres proteïnes dins la subfamilia, son DI1, DI2 i DI3. Per a estudiar aquestes proteïnes hem usat com a seqüència de referència la de Bos Taurus (vaca), això ens permetrà tenir un genoma més similar al de Bos grunninens Mutus que no el d’humà. D’altre banda hem de tenir en compte que el genoma humà està molt més estudiat i per tant l’anotació és més bona.
|
| |
Família Selenofosfat Sintases (SPS)a
La selenofosfat sintetasa és un enzim que s’encarrega d’afegir l’aminoàcid selenocisteïna a les selenoproteïnes durant la traducció. Aquest enzim té dues formes: SPS1 i SPS2. La primera no presenta selenocisteïnes i s’encarrega de reciclar la Sec i recuperar el seleni, mentre que la segona és la selenoproteïna homòloga en mamífers a Sel D en procariotes, i intervé directament en la traducció de les selenoproteïnes. Al genoma de Bos grunniens taurus hem trobat la SPS2. A partir del seleni, aquesta selenoproteïna participa en la síntesi de monoselenofosfat, necessari per la producció de la selenocisteïna.
|
| |
Família Tioredoxina Reductases (TR)a
Aquesta família de proteïnes representen els únics enzims capaços de reduir la Tioreduxina (Trx); concretament, catalitzen la reducció del disulfur del centre catalític de la tioredoxina per tornar–la activa. Cada monòmer de la proteïna conté un grup prostètic FAD i un domini d’unió a NADPH. És important en la regulació de l’ambient redox intracel·lular (defensa davant de processos d’estrès oxidatiu). TR1 es troba fonamentalment al citosol, TR2 a nivell mitocondrial i TR3 és específica dels testicles.
|
|
| |
| Selenoproteïnes en eucariotes |
|
| |
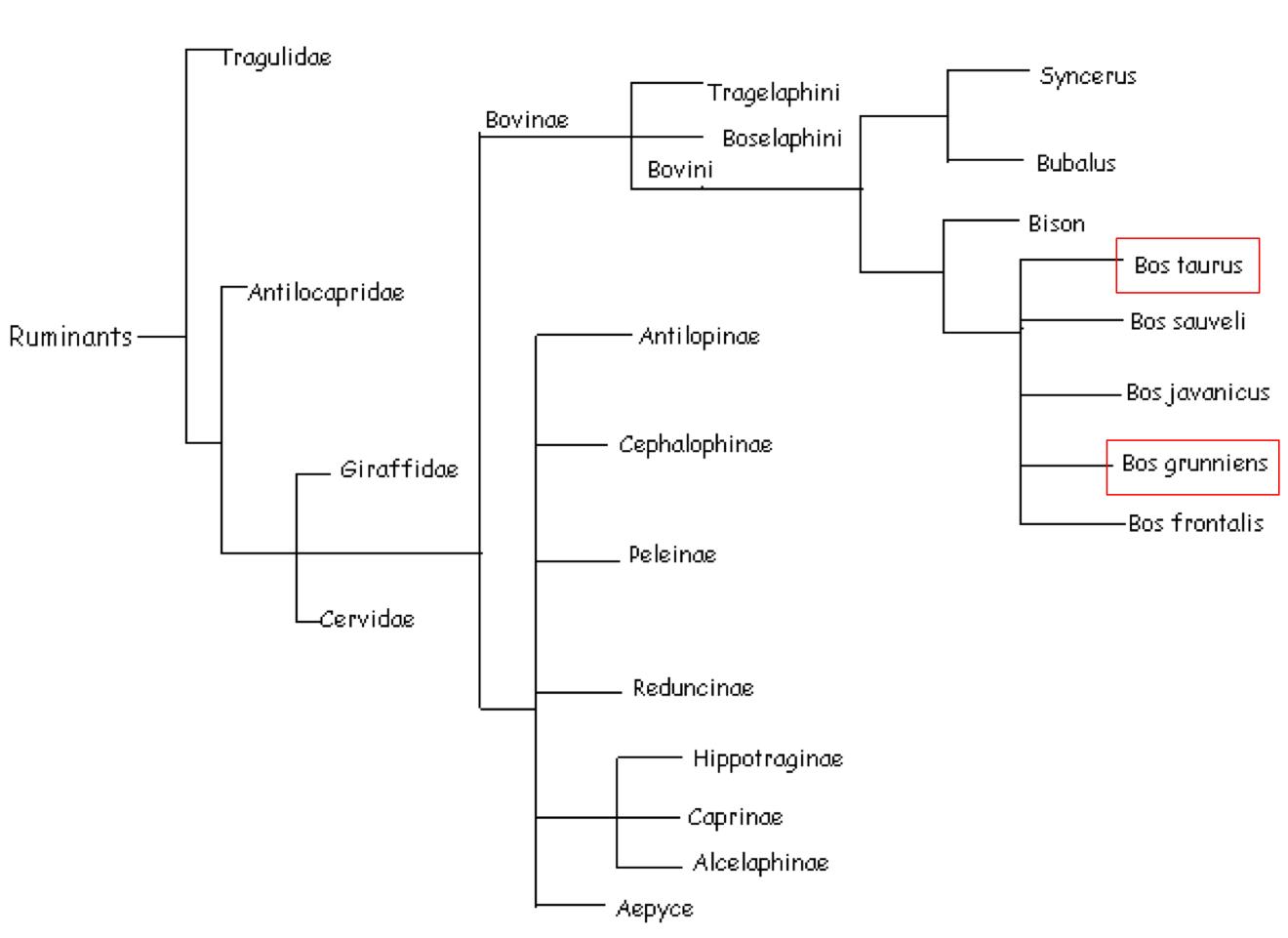
Aquesta imatge il·lustra les selenoproteïnes presents en el genoma dels organismes vertebrats. Com es pot veure, a més complexitat de l’organisme, més nombre de selenoproteïnes. Les que estan escrites en color vermell són les selenoproteïnes ancestrals (les subratllades són les que només s’han trobat en vertebrats), mentre que els verdes són les que han estat creades de nou, probablement per duplicacions, les grises les que s’han perdut i les blaves les que han patit una substitució d’una selenocisteïna per una cisteïna.
Segons aquest arbre, les selenoproteïnes comunes en tots els vertebrats són: GPx1, GPx2, GPx3, GPx4, TR1, TR3, DI1, DI2, DI3, SelH, SelI, SelM, SelN, SelO, SelP, MsrB1, SelS, SelT1, SelW1 i Sep15.
|
|
| |
|
|
| |
Puja dalt |
|
| |
|
|
| |
Classifiació taxonòmica
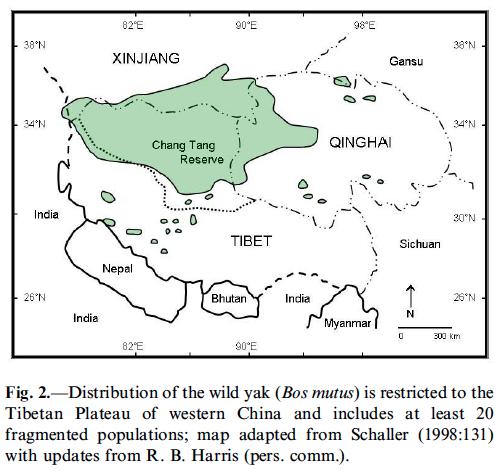
El Iac ( Bos grunniens mutus ) és una espècie bovina de pelatge llanós i mida mitjana que habita a l’Himàlaia, les muntanyes de l’Àsia Central, l’altiplà del Tibet, Mongòlia i Rússia. Pot arribar a viure als 6000m sobre el nivell del mar, s’alimenta de plantes baixes i no té depredadors (tot i que pot patir atacs d’óssos entre d’altres).
La paraula Iac prové de l’idioma tibetà i només s’utilitza per referir–se als mascles; les femelles es coneixen com a Dri o Nak. És capaç d’arribar fins als 20 anys de vida i acostuma a pesar entre 200 i 400 kg. Pot ser tan salvatge com domèstic; en aquest últim cas se n’utilitza la llana, la pell, la carn i la llet, de la qual se’n fan formatges o iogurts de gust molt fort. Per altra banda, el Iac salvatge, de major mida que el domèstic, habita només al Tibet i és una espècie amenaçada.
L’animal presenta una gepa i unes banyes llargues i cargolades que poden arribar a 1m de longitud. Malgrat s’han intentat fer anàlisis del seu ADN mitocondrial per determinar la seva història evolutiva, aquests han resultat ser una mica confusos. Aparentment podria haver divergit del toro fa entre 1 i 5 milions d’anys, tot i que tot indica que podria estar més estretament relacionat amb el bisó que amb els altres membres del seu propi gènere.
Encara no s’han identificat les selenoproteïnes presents a l’espècie Bos grunniens mutus , però sí que han estat caracteritzades les que hi ha a Bos taurus (vaca), espècie que es troba propera filogenèticament a la que s’estudia en aquest cas. És per això que s’ha escollit Bos taurus per buscar fer l’alineament de selenoproteïnes entre aquesta espècie i la d’interès i mirar si estan conservades.
|
|
| |
| | |