|
||||||
| Resum | Introducció | Materials i Mètodes | Resultats | Discussió | Conclusions | Referències |
|
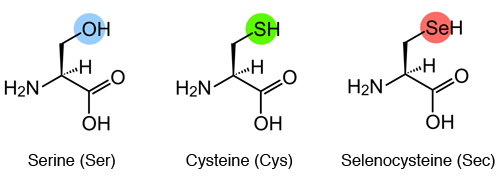
IntroduccióEl seleni és un micronutrient essencial per la vida de molts organismes. En humans, tant l’excés com el defecte d’aquest nutrient pot donar lloc a patologies. Amb el dèficit de seleni apareix la malaltia de Keshan que té una afectació cardíaca mentre que l’excés d’aquest pot donar lloc a intoxicacions. A la natura, el seleni no es troba en forma lliure sinó que, per evitar la seva toxicitat, forma part de l’aminoàcid selenocisteïna, present en les selenoproteïnes. [1] SelenocisteïnesLa selenocisteïna és l’aminoàcid número 21, està codificat pel codó TGA i es representa amb les abreviatures Sec i U. Estructuralment la selenocisteïna i la cisteïna són molt semblants però en la selenocisteïna trobem un àtom de seleni (grup SeH) substituint el grup tiol (grup SH) de la cisteïna. La semblança funcional entre selenocisteïna i cisteïna fa que, degut a processos evolutius, es puguin trobar proteïnes amb seqüències ortòlogues que tinguin la selenocisteïna substituïda per una cisteïna. [2]
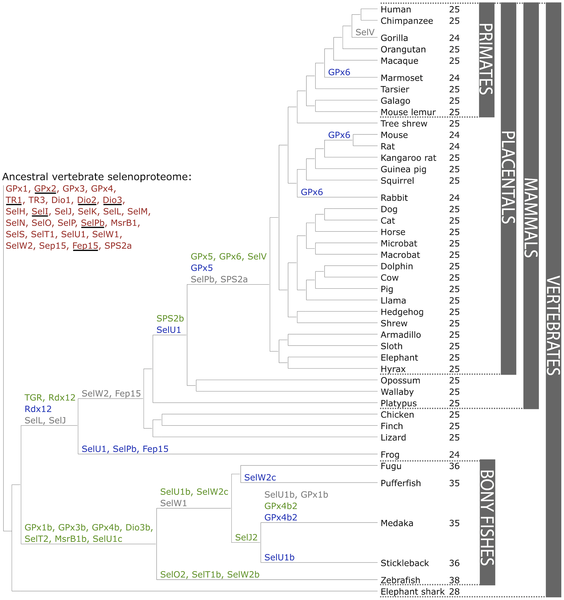
Selenproteïnes i la seva filogèniaLes selenoproteïnes són, en la seva majoria, enzims que presenten una selenocisteïna en el seu centre actiu. Hi ha una gran quantitat i varietat de selenoproteïnes distribuïdes en els tres dominis de la vida: bacteris, Archaea i eucariotes, però NO en les plantes. A més a més, s’ha observat que hi ha animals que no en presenten. Fins al moment s’han descrit 45 famílies de selenoproteïnes. En peixos podem trobar fins a 41 d’aquestes famílies i en mamífers fins a 28 malgrat que un sol mamífer pot tenir fins a 25 selenoproteïnes en el seu genoma. [3] A grans trets, les selenoproteïnes funcionen com a enzims redox i es creu que podrien tenir capacitat antioxidant, antitumoral i immunològica a més de funcionar com a quelants del seleni per disminuir-ne la toxicitat. No obstant això, es desconeix la funció d’algunes famílies de selenoproteïnes i la distribució de les selenoproteïnes al llarg de la línia evolutiva és incompleta. [4]
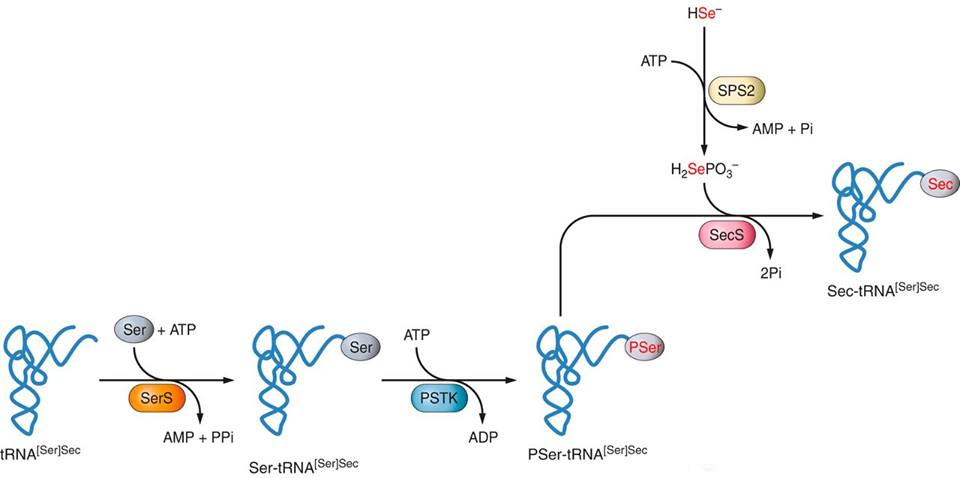
Síntesi de selenoproteïnesLa maquinària que s’encarrega de regular el procés de síntesi de selenocisteïnes i selenoproteïnes està formada pels següents enzims: SPS1,SPS2, SecS, Pstk, eEFsec i SBP2. [5] Síntesi de selenocisteïnaL’única forma de seleni capaç de formar part de les selenocisteïnes és el selenofosfat (H2PO4SeH) obtingut gràcies a la modificació que duu a terme l’enzim Selenofosfat Sintetasa (SPS2) del seleni ingerit per la dieta. L’estructura encarregada d’incorporar la selenocisteïna a la cadena d’aminoàcids en formació durant la traducció de la proteïna és el selenocisteinil-tRNA (tRNA +selenocisteïna), també anomenat tRNA[Ser]Sec. La selenocisteïna és l’únic aminoàcid que se sintetitza directament al tRNA i ho fa de la següent manera:
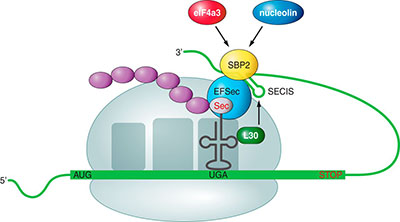
Incorporació de la selenocisteïna durant la traduccióHabitualment, el codó UGA (corresponent al DNA TGA) és un codó STOP però en les seqüències codificants per una selenoproteïna, la maquinària transcripcional no l’interpreta com un STOP sinó que hi incorpora la selenocisteïna i segueix la traducció fins al següent codó STOP. [5][6] En eucariotes, el mecanisme mitjançant el qual aquest codó s’interpreta de manera diferent esta mediat per la presència a l’extrem 3’ UTR d’una estructura tridimensional en forma de loop de RNA anomenada SECIS (Selenocystein Insercion Sequence). Aquesta permet la incorporació de la selenocisteïna a la cadena d’aminoàcids en formació gràcies al mecanisme següent:
Famílies de selenoproteïnesDILes Iodotironines deiodinases (DI) són un grup de selenoproteïnes homodimèriques que estan relacionades amb el metabolisme de l’hormona tiroidea. Catalitzen la transformació de la pro-horomona T4, generant isòmers actius de triiodotironina (T3), i inactius de triiodotironina reversa (rT3) i diiodotironina (T2). Aquesta biotransformació de l’hormona tiroidea es dóna pràcticament en tots els teixits i és catalitzada per tres isotips desiodinases diferents. DI1 (fetge i ronyo), DI2 (cor, múscul esquelètic, sistema nerviós central, teixit adipós, tiroides i glàndula pituitària) i DI3 (teixit fetal i placenta). Les deiodinases DI1 i DI2 serveixen per activar o desiodar l’anell exterior (ORD) de l’hormona. I la DI3 catalitza principalment la desactivació o iodació de l’anell interior de la pro-hormona T4 i T3 a rT3 i T2, respectivament. [7] GPxEls enzims glutatió peroxidases constitueixen una família de 8 selenoproteïnes en mamífers importants en el sistema del sistema cel·lular de defensa antioxidant i tenen una alta conservació. La funció principal de les glutatió peroxidases és protegir l’organisme de l’efecte oxidatiu i degradant dels peròxids i hidroperòxids formats de forma endògena. Aquestes proteïnes tenen una selenocisteïna present al lloc catalític, i la disposició de seleni regula l’enzim glutatió peroxidasa. [8] Cada GPX té una localització i una funció específiques:
MsrA i SelREls pèptids metionin sulfòxid reductases, MsrA i MsrB (SelR) no comparteixen homologia ni en l’estructura ni en la seqüència i, per tant, tenen un origen evolutiu diferent. Es tracta de pèptids que tenen una funció reparadora de proteïnes que han estat inactivades per oxidació en metionines. Catalitza la reducció de les metionines-sulfòxid a metionines. De fet, en mamífers MsrA és l’única proteïna capaç de dur a terme aquesta funció (reduir el metionenin-S-sulfòxid). En mamífers MsrA conté una cisteïna, i MsrB una selenocisteïna. Així mateix, MsrA en procariotes conté una selenocisteïna. MsrB presenta tres isoformes: SelR1, SelR2 i SelR3, entre les quals SelR3 i SelR2 tenen major homologia. [9] SelHSelH és una proteïna oxidoreductasa thioredoxin-like nuclear amb un motiu CXXU conservat (Cisteïna seprada per dos residus qualsevols d’una selenocisteïna). Aquest motiu suggereix una funció redox de SelH i també ens indica que aquesta depèn de l’element traça seleni. No se’n coneix homologia amb altres selenoproteïnes caracteritzades. Aquesta estructura també és present en varies selenoproteïnes en mamífers (SelW, SelT, SelM). [10] Sel ILa Selenoproteïna I és una proteïna que conté una selenocisteïna prop de la regió C-terminal. Se sap que té un domini CDP-alcohol fosfatidiltransferasa molt conservat que típicament es troba en les colina-fosfotransferases (CHPT1) i les colina fosfotransferases/etanolamina fosfotransferases (CEPT1). CHPT1 catalitza la transferència d’una colina a diacilglicerol des de CDP-colina i CEPT catalitza una reacció semblant però accepta tant colina com etanolamina. Sel I catalitza la biosínesi de fosfatidiletanolamina a partir de CDP-etanolamina. Aquesta selenoproteïna té predits 7 dominis transmembrana (similars als de CHPT1 i CEPT1). Una seqüència important de la proteïna es troba entre el primer i el segon dominis transmembrana, on hi ha localitzats tres residus d’àcid aspàrtic crítics per la seva funció. [11] Sel KLa selenoproteïna K és una proteïna transmembrana localitzada al reticle endoplasmàtic amb un nivell d’expressió elevat en determinats teixits, com poden ser el cor, la melsa, algunes regions cerebrals i les cèl·lules immunes. Es troba en moltes espècies d’eucariotes superiors i molts estudis suggereixen que SelK està involucrada en la regulació del calci i en la via de degradació proteica associada al Reticle endoplasmàtic. Aquesta via de degradació proteica consisteix en un sistema responsable de l’eliminació de proteïnes en el citoplasma si aquestes no s’han plegat correctament en el RE. Es creu que SelK té una estructura ben definida perquè té dominis SH3 i és rica en Glicines, Prolines i residus amb càrrega. S’està estudiant si la seva funció té a veure amb una resposta a l’estrès oxidatiu del reticle endoplasmàtic. [11] Sel MLa Selenoprotein M és una proteïna que es troba en el reticle endoplasmàtic i te dues regions d’interès: un motiu redox CXXU i un plegament de tipus thioredoxin-like (TRX). Té un paper rellevant en la regulació del calci i la protecció enfront l’estrès oxidatiu. S’expressa principalment al cervell. En moltes espècies de vertebrats aquesta selenoproteïna s’ha perdut. Els residus terminals semblen tenir un signe KDAL de retenció en el reticle endoplasmàtic ja que és perinuclear. SelM és un homòleg llunyà de Sel15. Es localitza en el RE/Golgi on s’associa amb la UDP-glucosa (glicoproteïna glucosiltransferasa), una proteïna resident al RE involucrada en el control de qualitat del plegament proteic; controla si els enllaços disulfur estan ben fets. [5] Sel NLa selenoproteïna N és una glicoproteina del reticle endoplasmàtic amb un patró d’expressió del desenvolupament temprà. Té un paper fonamental en la funció muscular després del naixement; no obstant això, durant l’edat adulta es troba en molt baixes concentracions. Aquesta selenoptoreïna està formada per dues isoformes i s’expressen en el múscul esquelètic, el cervell, el pulmó i la placenta. La isoforma II també s’expressa en cor i l’estómac. Defectes en l’expressió del gen que codifica per SelN (SEPN1) donen lloc a distròfia congènita muscular amb rigidesa espinal (RSMD1). [12] Sel OLa Selenoproteïna O és una proteïna que està àmpliament distribuïda: té homologies en animals, bacteris, llevats i plantes. SelO adopta un plegament tridimensional semblant al de les kinases, sense conservar el residu d’apartat en el domini catalític His-Arg-Asp. Possiblement aquest domini quinasa li dóna un paper en la xarxa de senyalització cel·lular. En llevats i bacteris té un paper en la resposta d’estrés oxidatiu. També s’ha vist un paper important en el transport ABC i en la regulació del metabolisme. Només homòlegs de SelO en vertebrats tenen Selenocisteïnes, les quals estan localitzades en la penúltima posició de l’extrem C-terminal. Al igual que la SelM, el loop apical de l’element SECIS hi ha dues citosines enlloc de dues adenines (vist en mamífers). [13] Sel PLa selenoproteïna P té múltiples residus de selenocisteïnes - 10 en humans - localitzats en el domini N-terminal (domini enzimàtic) i C-terminal (domini de transport) i és la selenoproteïna més comuna al plasma. És indispensable per l’homeòstasi del seleni en l’organisme i promou la retenció d’aquest en el cos i afecta el transport de seleni del fetge als teixits extra-hepàtics. La SelP també pot veure’s implicada en el metabolisme de la glucosa regulant la secreció d’insulina en el pàncrees. A més, té un poder antioxidant gràcies a la cpacitat d’unir heparina i metalls pesant presents en l’organisme com el cadmi i el mercuri. El domini C-terminal presenta 9 Selenocisteïnes i el domini N-terminal en presenta 1. S’ha relacionat la SelP amb la malaltia de l’Alzehimer perquè s’ha vist colocalització de SelP amb les plaques d’amiloide. [14] Sel SLa Selenoproteïna S pot tenir presència de SECIS o no segons l’splicing que pateixi el seu RNA druant la transcripció. L’expressió de SelS està associaciada a malaltia cardiovascular en diabetes i està regulada com a resposta cel·lular a glucosa i insulina. A més, també s’ha vist associada a la regulació de la resposta inflamatòria i, per tant, regulada per citokines inflamatòries. En la membrana del recicle endoplasmàtic participa en la retrotranslocació de proteïnes mal plegades cap al citsol, on seran degradades. Té a veure amb la resposta a estés oxidatiu. [15] Sel TLa selenoproteïna T és una selenoproteïna petita amb un motiu putatiu redox N-terminal. S’intueix que la seva localització és a la membrana plasmàtica per la presència d’un domini hidrofòbic. [16] Sel ULa Selenoproteina U és una selenoproteïna que trobem en peixos i molt altres organismes marins, ocells i algues; el seu homòleg en cisteína, és present en mamífers, artròpodes, cucs, amfibis i plantes terrestres. En mamífers s’han predit tres isoformes diferents: SelU1, SelU2 i SelU3. Actualment, se’n desconeix la funció. SelU1 s’expressa en teixits com l’òs, el cervell, el fetge i el ronyó. [17] Sel WLa Selenoproteïna W és la selenoproteïna més petita dels mamífers. S’expressa en diferents organismes eucariotes i en bacteries, i la seva distribució no es restringeix a un únic òrgan en els mamífers. S’ha vist que piodria tenir un paper en la regulació de la mutagènesi, del tracte gastrointestinal i del sistema inmunitari. [18] TRLes Thioredoxin reductases són proteïnes amb seqüència semblant a les glutatió reductases. Tenen un domini redox catalític consevat amb: Cy-Val-Asn-Val-Gly-Cys-. La seva funció és la de reduir les tiroreduccines. Els mamífers tenen tres isoenzims de TR; el TR1 que es troba al citosol i el TR2 i el TR3 que tenen localització mitocondrial. [19] Balaenoptera acutorostrata
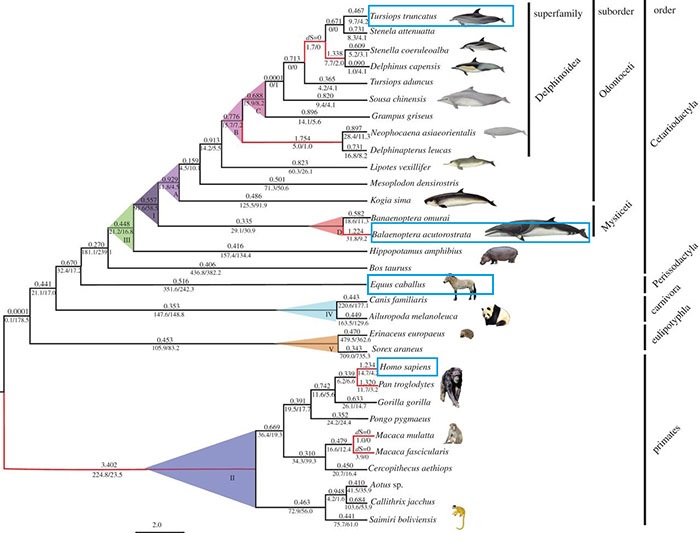
La Balenoptera acutorostrata, anomenada comunament rorqual d’aleta blanca, és un mamífer de grans dimensions que habita a les aigües de grans oceans i mars de tot el món. És l’espècie més petita de rorquals i pot arribar a mesurar 10,2 metres de llarg i pesar 10.000kg. Malgrat no ser considerats costaners, aquests rorquals no s’allunyen més de 169km de la costa i a vegades entren a estuaris, badies, fiords i llacs. La seva distribució és extensa i variada al llarg dels diversos oceans, però prefereix regions fredes i polars a regions més temperades i tropicals. Igual que la majoria de balenes, aquest rorqual s’alimenta principalment de krill i d’alguns peixos petits però la seva dieta es torna més omnívora com més al nord es localitzen. Habitualment es desplacen sols o en petits grups (2-4) però aquest poden augmentar en les zones més riques en krill. La gestació dura 10 o 11 mesos i només tenen una cria per cada embaràs. Les cries pesen 450kg al néixer i s’alimenten de llet materna durant 5 mesos. No obstant això, no assoleixen la maduresa sexual fins als 6 anys. [20]
Tria dels genomes de referènciaCom ja s’ha comentat anteriorment, l’espècie Balaenoptera acutorostrata pertany a l’ordre Cetacea. És per això, que s’ha comparat el genoma problema amb les proteïnes homòlogues del dofí Tursiops truncatus, una espècie que també pertany a l’ordre Cetacea. A més a més, l’anàlisi també es va dur a terme comparant amb les d'Homo sapiens. Aquest és el genoma més estudiat i, per això, ens pot permetre determinar si alguna de les selenoproteïnes no trobades al comparar amb dofí la trobem al comparar amb humà. Per últim, també s'ha triat l'espècie Equus caballus per l'anàlisi de determinades proteïnes on el resultat al comparar en dofí i humà no era clar. Aquesta espècie de cavall és relativament propera al rorqual en la línia filogenètica.
|
| Components del grup: Mar Costa, Anna González, Clara Mayayo, Isshak Mrabet, Mirna Muntal |