Selenoproteïnes
en Elephantulus edwardii
Brasó M, Güell N, León S, López A
El Seleni
El seleni és un micronutrient essencial per a la vida animal la principal font d'obtenció del qual és la dieta. El seleni és conegut principalment per les seves propietats antioxidants així com per la seva implicació en la funció immunitària i el metabolisme de les hormes tiroidees. A més, a nivell més terapèutic es caracteritza per les seves propietats antiinflamatòries i antivirals. Per tant, la deficiència d'aquest aminoàcid estarà relacionada amb problemes de salut, entre d'altres, la malaltia de Keshan [7], [8], [12].
Com a curiositat, el seleni va ser descobert pel químic suec Jöns Jacob Berzelius, qui va posar-li el nom de Selene en honor a la deesa grega de la lluna [9].
Les Selenoproteïnes
Les selenoproteïnes són un tipus de proteïnes que contenen un aminoàcid anomenat selenocisteïna (Sec) en la seva estructura. L'estructura molecular de la selenocisteïna consisteix en un anàleg de l'aminoàcid cisteïna en el qual s'ha substituït el àtom de sulfur per un àtom de seleni [1] [3].
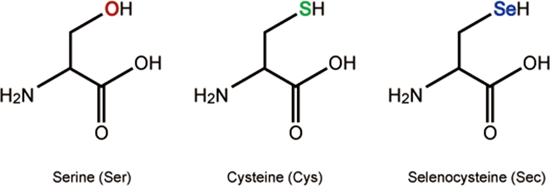
La selenocisteïna no és un dels vint aminoàcids clàssicament estudiats, i per tant, no es troba en el codi genètic estàndard. El codó que codifica les Selenoproteïnes és el UGA, que resulta ser un Stop Codon. De manera que, per tal de sintetitzar selenoproteïnes caldrà maquinària específica que recodifiqui aquest codó per tal que no s’aturi la traducció [5].
Biosíntesi de les selenoproteïnes
El procés de síntesi de les selenoproteïnes ha estat conservat evolutivament, tot i que els mecanismes són diferents entre els diferents regnes. Tot i les diferències específiques, els elements en comú són el codó UGA-Sec, el tRNA específic per a la síntesi, l'element SECI i alguns factors proteics [7].
Per dur a terme la incorporació del seleni en forma de selenocisteïna a les selenoproteïnes, tal i com s'ha dit en l'apartat anterior, cal que es recodifiqui el codó stop UGA. L’element que recodifica el codó UGA és una estructura secundària de RNA anomenada SECI (SelenoCysteine Insertion Sequence) situada en l'extrem 3'UTR del mRNA [7].
L'element SECI és l'encarregat d'unir-se a SBP2 o SECIs Binding protein, la qual reclutarà el factor d'elongació eEFSec, que al seu temps, s'uneix al tRNA específic per la traducció de selenoproteïnes, unit a l'aminoàcid selenocisteïna. Una vegada es compleixin aquestes condicions, la traducció continua fins el pròxim codó STOP [7].
Tipus de selenoproteïnes
Les selenoproteïnes es troben distribuïdes en els tres regnes: bacteris, Archaea i eucariotes, tot i que no en totes les espècies d'aquests tres regnes [4].
La mida dels selenoproteomes varia de manera significant entre els eucariotes, de manera que el contingut més alt en selenoproteïnes es troba en organismes aquàtics [3].
S’han decotbert 25 selenoproteïnes en mamífers 6 . Les selenoproteïnes dels mamífers es classifiquen en dos grups segons la posició de la selenocisteïna; si aquesta és pròxima al C-terminal, el grup consta de la TrxRs, i les selenoproteïnes S,R,O, I i K, mentre que si la selenociteïna està a l’N-terminal, l’estructura és com la de la tioredoxina, ja que contenen el domini CXXU, que correspon al domini actiu de les tioredoxines. Per la seva estructura, la funció de moltes selenoproteïnes està relacionada amb reaccions redox. Tot i així, la funció exacta de moltes selenoproteïnes, és encara desconeguda. Seguidament s’explicaran breument les característiques principals de cada selenoproteïna [3], [7] i [9].
A més, no totes les selenoproteïnes es troben en tots els organismes; per exemple, la SelU és una proteïna que es troba en peixos (com el fugu) però no en humans. És important destacar que a vegades la seqüència ortòloga d’una selenoproteïna conté una cisteïna en comptes d’una selenocisteïna [7], [8] .
- Tioredoxin Reductases (TrxR)
- Glutatió peroxidades (GPx)
- Iodotironin deiodinases (DIOs)
- SPS
- Selenoproteïna 15
- Selenoproteïna H
- Selenoproteïna I
- Selenoproteïna K
- Selenoproteïna M
- Selenoproteïna N
- Selenoproteïna O
- Selenoproteïna P
- Selenoproteïna W
- Selenoproteïna R
- Selenoproteïna S
- Selenoproteïna T
- Selenoproteïna V
Les tioredoxin reductases i la tioredoxina (Trx), juntament amb el NADPH, constitueixen el sistema redox de les cèl·lules. Aquests enzims, de 115 kDa aproximadament, tenen una gran homologia amb les glutatió reductases, tot i que el seu mecanisme de catalització és diferent. Són proteïnes molt versàtils, ja que poden actuar controlant la funció redox la molècula tioredoxina o bé com a reductors de diferents substrats. La Trx és l’element central de la regulació redox mitjançant la seva capacitat reductora de disulfit i és el substrat principal de les TrxR. Té una funció imprescindible ja que catalitza la reducció de la ribonucleotid reductasa, enzim necessari per a la síntesi de DNA. S’han identificat 3 selenoproteïnes TrxR en mamífers: TrxR1 al citosol, TrxR2 al mitocondri i TrxR3/tioredoxin-glutatió reductasa al testicles. A més a més, es pot donar splicing alternatiu en el gens que codifiquen per aquests enxzims, de manera que és possible que hi hagi una gran varietat d’espècies de TrxR. [4], [7].
Van ser la primera família de proteïnes descobertes d’introduir seleni al domini catalític i és per això que va ser relacionat amb la funció antioxidant del seleni. La família de les glutatió peroxidases són conegudes per actuar com antioxidants catalitzar la reducció del peròxid d’hidrogen (H2O2) entre d’altres, protegint les cèl·lules del dany oxidatiu. Es coneixen 7 GPx en humans, dues de les quals contenen cisteïna en comptes de seleni [7].
Gpx1, present al citosol, actua com a antioxidant reduint directament H2O2; Gpx2 específic del sistema gastrointestinal, que protegeix davant els hidroperòxids lipídics ingerits i té una gran importància al colon, Gpx3, és secretada al plasma i té una gran varietat de substrats, fent-la molt eficient en la funció antioxidant al plasma. Gpx4 està present al citosol i al mitocondri en diferents tipus cel·lulars. Pot actuar sobre molts substrats i té especial importància en la maduració de l’esperma, actuant com a promotor de la maduració d’aquest. Gpx6 es troba a l’epiteli olfactori i als teixits embrionaris [4], [7] i [9].
Pel que fa a les homòlogues amb cisteïna, Gpx5 és específica de l’epidídim i Gpx7 correspon a la fosfolípid hiperòxid glutatió peroxidasa [7].
Corresponen a la família d’enzims relacionats amb les hormones tiroidees, ja que en catalitzen la reacció d’activació o inactivació. Com són hormones amb una gran importància, els patrons d’expressió de les DIOs són diferents al llarg del desenvolupament i als teixits humans adults. La DIO1 s’expressa bàsicament al fetge, la ronyó, a la glàndula tiroides i a la pituïtària i té un paper molt important en la producció de 3,3’5-triiodotironina (T3) ja que catalitza la deiodinació de la prohormona T4 a la glàndula tiroides i a més, regula la circulació de la mateixa.
Pel que fa a la DIO2, s’expressa a la glàndula tiroides, al SNC i a la glàndula pituïtària, al múscul esquelètic i al teixit adipós bru i, fa la mateixa funció que la DIO1.
La DIO3, en canvi, s’expressa bàsicament als teixits fetals (placenta, fetge, cervell i pell) i a l’úter de la mare i converteix la T4 en la T3 reversa [4], [7] i [9]. A més, la funció d’aquestes dues és a nivell de processos teixit i òrgan específics [7].
Les selenofosfat sintetases són selenoproteïnes essencials per a la síntesi de les selenoproteïnes incloent la seva pròpia ja que està involucrada en la biosíntesi de la selenocisteïna. Catalitzen la conversió de selenit en selenofosfat, el donador de seleni necessari per a la formació de Sec [7].
Aquesta selenoproteïna s’expressa a molts teixits, destacant-ne nivells elevat al cervell, pulmons, fetge, glàndula tiroides, ronyons i testicles. Tot i que la seva funció no està del tot clara, es pensa que, donada la seva localització al reticle endoplasmàtic, intervé en el correcte plegament de les gicoproteïnes [7].
La SelH es troba localitzada al nucli i està àmpliament distribuïda per molts teixits i s’expressa als estadis primerencs del desenvolupament embrionari. És una proteïna amb plegament similar a las tioredoxines fet que suggereix que la seva funció consisteix en regular l’expressió gènica en la síntesi de glutatió i a més a més, juga un paper en la resposta a les reaccions redox [8].
Aquesta selenoproteïna transmembrana s’expressa en tots els teixits en els quals es dóna la síntesi de fosfolípids, ja que conté una seqüència homòloga als enzims encarregats de la síntesi d’aquestes molècules [8]. .
La selenoproteïna K, present dins el RE i a la membrana plasmàtica, s’expressa principalment al múscul esquelètic i al cor, tot i que altres teixits com el pàncrees, el fetge i la placenta també presenten expressen SelK. Actua com a antioxidant ja que disminueix els nivells de ROS intracel·lular i protegeix el cardiomiòcits de l’estrés oxidatiu [7].
La SelM s’expressa al cor, als pulmons, als ronyons, a l’úter i la placenta i en grans quantitats a la tiroides i al cervell. Està implicada en reaccions redox. És interessant el fet que el gen responsable de l’aparició d’Alzheimer inhibeix la SelM [7].
La SelN, expressada a la membrana del reticle endoplasmàtic dels teixits, és la única selenoproteïna directament relacionada amb malaltia. Mutacions que codifiquen pel gen d’aquesta (SEPN1) i al seu element SECIS són causants de miopaties [7] .
Tot i que es sap que la SelO consta de 669 aminoàcids, la seva localització, distribució i funció es desconeixen. No obstant, conté un domini que suggereix que la seva funció està relacionada amb les reaccions redox [8].
La selenoproteïna P és secretada al plasma pel fetge en la seva forma glicosilada i s’expressa a tots els teixits. És la segona més present en plasma després de la GPX3. La seva funció consisteix en transportar el seleni als teixits, ja que està associada a la membrana cel·lular [4], [7] i [9].
És una selenoproteïna altament conservada en mamífers que s’expressa als teixits en funció de la quantitat de seleni a l’organisme. No obstant., el seu nivell al cervell és constant. Durant el desenvolupament embrionari s’expressa a la implantació i gastrulació i, conseqüentment, al sistema nerviós, pulmons i cor. Es localitza principalment al citoplasma i una petita fracció unida a la membrana cel·lular. Com que la SelW uneix glutatió amb una gran afinitat, es creu que la seva funció és actuar d’antioxidant 7. Tot i això, també s’ha vist que està expressada als progenitors musculars, de manera que probablement tingui un paper essencial pel múscul i el seu metabolisme. És per això que es creu que està implicada malalties musculars [4], [7] i [9].
Aquesta selenoproteïna, també coneguda com a metionin-R-solfoxid reducasa, catalitza la reducció dels residus oxidats de metionina obtinguts en resposta a l’increment d’elements ROS. És per això que la seva activitat redox és necessària per la protecció de les proteïnes danyades [7] .
L’expressió d’aquesta selenoproteïna és induïda en resposta a l’estrés del reticle endoplasmàtic (RE), mentre que és inhibida per la glucosa i a més a més, interactua amb el sèrum amiloide A, una proteïna inflamatòria de fase aguda. És per això que es creu que està relacionada amb la Diabetes Mellitus tipus 2, la inflamació i la malaltia cardiovascular i té un paper important en el control de la resposta inflamatòria [7] .
La selenoproteïna T es localitza al RE i la seva seqüència i plegament és similar al de las tioredoxines. Donada la seva expressió, es creu que la seva funció està relacionada amb els fenòmens d’estrés cel·lular i en l’activació del polipèptid activador de l’adenilat ciclasa a la hipòfisi mitjançant la regulació del calci [8].
L’expressió d’aquesta selenoproteïna està restringida als testicles però la seva funció és encara desconeguda. Tot i així, la seqüència de la SelV és homòloga amb altres selenoproteïnes amb plegament com les tioredoxines, de manera que es creu que està involucrada en funcions redox [8].
Les musaranyes Elefantulus d'edwardii són rosegadors del gènere Macroscelides, de la família monofilètica dels Macroscelididae. Tot i així, antigament estaven inclosos dins la família dels insectívors. Són originaris de Sud àfrica, formant part dels mamífers endèmics africans [2].
El nom de les musaranyes elefant prové de la semblança amb la trompa dels elefants, ja que tenen un musell allargat i móbil [13].

La massa corporal dels integrants del gènere dels Macroscelides oscil·la entre 35-50g, és a dir, són animals petits comparats amb altres espècies de la subfamília de Macroscelidinae [2].
Els Macroscelides tenen el crani gran a causa de la gran cavitat auditiva, unes orelles petites i arrodonides i un pelatge relativament llarg, dens i suau. De manera similar a altres gèneres dels Elephantulus, els Macroscelides tenen uns ulls grans, un musell molt llarg i flexible, una cua com la dels ratolins i unes potes llargues i ràpides. Tenen poc dimorfisme sexual pel que fa a les diferències en les mesures corporals[2].
 Són animals nocturns amb una dieta omnívora principalment de preses invertebrades[2] . Tot i així, recentment s’ha vist que també s’alimenten de nèctar de determinades espècies de plantes, com per exemple, de la Whiteheadia bifolia [13] .
Són animals nocturns amb una dieta omnívora principalment de preses invertebrades[2] . Tot i així, recentment s’ha vist que també s’alimenten de nèctar de determinades espècies de plantes, com per exemple, de la Whiteheadia bifolia [13] .
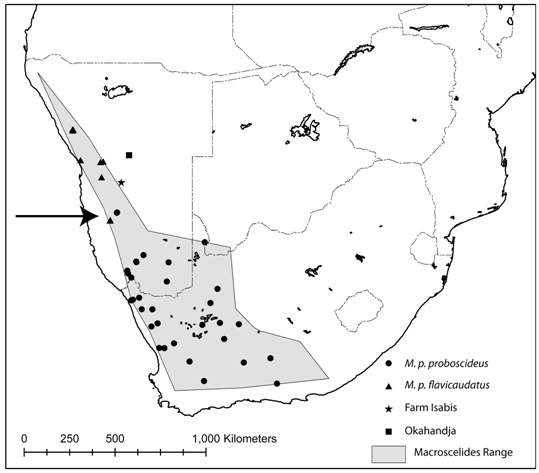
Els Macroscelides són una espècie monògama i pot ocupar un rang d’ocupació de fins a 1km2. Com altres membres de la subfamília dels Macroscelidinae, els Macroscelides no fabriquen nius sinó utilitzen coves a la vora dels matolls per deixar la progènie. Els Macroscelides es distribueixen des del nord-oest de Namíbia al sud per l'est d'àfrica del Sud [2].
Tot i que no és una espècie abundant, l’Elephantulus edwardii es troba en una àrea major a 130.000 km2, sent prou comú .
El seu hàbitat correspon a les zones rocoses i àrides, de manera que no es coneixen amenaces al llarg de Cape Sengi, sud àfrica. Es desconeix el nombre d’animals de la població actual, però no hi ha cap factor, més enllà de la variació natural de les condicions ambientals a les zones rocoses i àrides, per creure que l’espècie està augmentant o disminuint[2].
Filogènia
Tal i com es pot veure a l'arbre filogenètic, l'Elephantulus edwardii, pertany a l'ordre dels Macroselidea i és filogenèticament molt pròxim a l'ordre Rodentia. No obstant, tot i que tenen un ancestre comù, els Macroscelids van divergir abans que els Rodentia.
Com el genoma de Mus musculus (Rodentia Muridae Mus musculus) està molt ben caracteritzat ja que és el model animal d'estudi per excel·lència, es va decidir utilitzar el seu genoma per comparar-lo amb el d'Elephantulus edwardii i dur a terme la recerca de les selenoproteïnes.

Índex
- El seleni
- Les selenoproteines
- Biosïntesi
- Tipus de selenoproteïnes
- Elephantulus edwardii