CONCLUSIONS
Aquest apartat pretén englobar una conclusió general dels resultats obtinguts en la recerca de les dues famílies de selenoproteïnes, Thioredoxin reductase i Fep15, en els diferents genomes seqüenciats al 2011. L'objectiu és analitzar-los globalment per poder comprendre el significat d'aquests i entendre el comportament de l'evolució de les selenoproteïnes. Alhora, hem aprofitat aquest espai per remarcar les troballes més interessants.
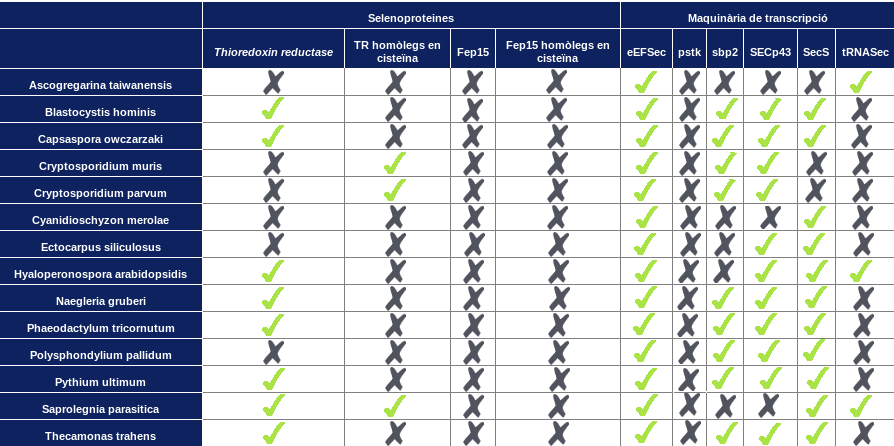
Primerament, hem sintetitzat els resultats obtinguts en una taula per poder-los veure d'una manera més esquemàtica:
Tot seguit, hem dut a terme un anàlisi global per a cada família de selenoproteïnes.
Després de dur a terme un anàlisis exhaustiu dels diferents genomes seqüenciats al 2011, han estat identificades nou selenoproteïnes de la família Thioredoxin reductase (TR) en vuit dels catorze protistes estudiats. Concretament, s'han trobat dues selenoproteïnes a Blastocystis hominis i una a Capsaspora owczarzaki, Cryptosporidium muris, Cryptosporidium parvum, Hyaloperonospora arabidopsidis, Naegleria gruberi, Phaeodactylum tricornutum, Pythium ultimum, Saprolegnia parasitica i Thecamonas trahens.
A més a més, també han estat identificats tres homòlegs en cisteïna en Cryptosporidium muris, Cryptosporidium parvum i Saprolegnia parasitica.
Així doncs, tenim un genoma, B.hominis, que conté dues selenoproteïnes i un altre genoma, S.parasitica, que conté una selenoproteïna i un homòleg en cisteïna de la mateixa família. Per tant, podem suposar que en aquests genomes s'ha produït una duplicació i que presenten, cadascun, dos paràlegs per la mateixa proteïna.
D'aquesta manera, es pot concloure que la família Thioredoxin reductase es troba molt estesa entre els diferents organismes estudiats, ja sigui com a selenoproteïna o com a homòleg en cisteïna. Això és degut a la importància de les funcions cel·lulars en què estan implicades les Thioredoxin reductases. Aquestes actuen en la reducció depenent de NADPH de les proteïnes Thioredoxins i d’altres components endògens o exògens subministrant equivalents reductors a diferents enzims i també s'ha vist que actuen com a factors de creixement cel·lular i d’inhibició de l’apoptosis. Així doncs, podem suposar que han de ser importants per al funcionament correcte de la majoria d'éssers vius.
Contràriament, no s'ha trobat cap selenoproteïna ni cap homòleg en cisteïna en els genomes d'Ascogregarina taiwanensis, Cyanidioschyzon merolae, Ectocarpus siliculosus i Pythium ultimum. Segurament aquests organismes supliran la funció d'aquestes amb altres proteïnes implicades en processos d'oxido-reducció. Hem observat que només dos d'aquests organismes es troben a la mateixa branca de l'arbre filogenètic dels protistes, pel que no podem concloure de manera clara que el fet que no presentin cap selenoproteïna ni cap homòleg en cisteïna de la família TR estigui relacionat evolutivament.
Per tal de determinar el grau de conservació entre totes les selenoproteïnes i homòlegs en cisteïna de la família de les Thioredoxin reductases que hem trobat en els diferents genomes, hem realitzat un alineament múltiple (veure) que ens ha permès determinar les regions més conservades entre aquestes i que el grau de similaritat que presenten en conjunt és força elevat.
També caldria remarcar que durant la recerca d'elements SECIS hem trobat un fet molt interessant en el genoma de P.tricornutum. En aquest organisme, que presenta selenoproteïnes de la família TR, no hem trobat elements SECIS a la regió 3' dels gens que codifiquen per aquestes selenoproteïnes, sinó que n'hem trobat a la regió 5'. Aquesta troballa és molt sorprenent ja que els organismes eucariotes es caracteritzen per presentar els elements SECIS a la regió 3' del gen que codifica per a la selenoproteïna corresponent. En canvi, els organismes del regne archaea es caracteritzen per presentar els elements SECIS a la regió 5' del gen. Així doncs, P.tricornutum podria estar representant un pont entre el regne archaea i el regne eucariota. No obstant, no podem afirmar-ho rotundament i hem de considerar que els elements SECIS trobats per a les dues selenoproteïnes de la família TR presents en aquest genoma puguin ser falsos positius.
Tal com suposàvem, després d'haver analitzat els diferents genomes, podem determinar que Fep15 segueix essent una selenoproteïna exclusiva de peixos ja que en cap dels protistes hem obtingut resultats.
Llavors, podríem hipotetitzar la història evolutiva de la selenoproteïna Fep15 com una aparició nova en la línia dels peixos o com una, tot i que més improbable, desaparició de la resta d'organismes. Per aquest motiu, seria necessari un estudi més exhaustiu i que comprengués les espècies més properes als peixos.
D'aquesta manera podria plantejar-se que la presència d'alts nivells de seleni al mar, podria ser un dels factors claus que hagués conduit a l'aparició d'aquesta selenoproteïna en els peixos. Això ho podríem estudiar investigant si es troba present en altres organismes marins i si l'aparició de noves selenoproteïnes és un procés afavorit. Si a més la seva funció ja està coberta en altres espècies per altres proteïnes (que no incorporen seleni), potser en els peixos l'aparició de Fep15 impliqui també la desaparició d'aquelles altres proteïnes que realitzaven la mateixa funció. Això es podria comprovar comparant amb altres organismes marins els genomes analitzats (tot i que s'hauria de tenir en compte molts factors).
La hipòtesi d’aquesta possible desaparició en la resta d'organismes també és plausible, tot i que menys probable, ja que implicaria que tingués lloc en moments diferents durant l'evolució i a més a més, en espècies molt allunyades entre elles (per exemple, els protistes que hem analitzat disten molt dels humans tant en la línia evolutiva com en les característiques que podrien haver-la fet desaparèixer).
Així doncs, si ens haguéssim de decantar, apostaríem per la primera opció i pressuposaríem que Fep15 és una selenoproteïna de nova aparició en els peixos, ja que tot i que no és coneguda, deu tenir una funció exclusiva en aquests animals.
Per estudiar quina pot ser aquesta possible funció, podríem realitzar un seguit d'experiments on observéssim diferents espècies de peixos en absència de Fep15 (per exemple fent knock-outs o en el cas que no fossin viables, amb siRNA).
També podríem, amb l'ajuda de programes informàtics, comparar-la amb altres selenoproteïnes de les quals en coneixem la funció dels seus dominis catalítics i veure si presenten conservació amb alguns d'aquests.
A més, com sabem que és un homòleg relativament distant de la família de Sep15 i la funció d'aquestes és realitzar ponts disulfur al reticle endoplasmàtic, podria ser que Fep15 tingués una funció similar ja que també es troba present al RE.
Tot i aquest gran nombre d'hipòtesis, cap l'hem pogut revocar o confirmar amb el treball que hem dut a terme. Senzillament, hem augmentat el número d'organismes que no presenten Fep15 limitant cada vegada més l'exclusivitat d'aquesta a les diferents espècies de peixos.