Introducció
Les selenoproteïnes estan presents en els tres dominis de la vida; bacteria, archea i eukarya. Els eucariotes tenen una distribució molt diferent de selenoproteïnes o selenoproteomes, que pot variar des de zero en plantes i fongs fins a més de trenta en alguns peixos i algues. En procariotes la distribució de selenoproteïnes també varia molt entre organismes. Els selenoproteomes d'espècies relacionades generalment són similars entre ells, però com més allunyades estan evolutivament, l'ús de selenoproteïnes es perd en varis fílums. A més alguns organismes han perdut la maquinària necessària durant l'evolució. En aquests organismes es poden trobar homòlegs de cisteïna (Cys) o arginina (Arg).
La inserció d'un Sec en una proteïna es determina pel codó UGA, però també necessita la conservació d'una estructura en forma de loop coneguda com a Sec insertion sequence (SECIS) element. En els eucariotes aquesta seqüència està localitzada a l'extrem 3' UTR del gen. La síntesis de selenoproteïnes és un procés conservat evolutivament, encara que s'han trobat grans diferències en el mecanisme de síntesis entre procariotes i eucariotes. Les característiques comunes a tots els organismes són la presència d'un codó UGA codificant per selenocisteïna, un tRNA específic, l'element SECIS i alguns factors proteics. Els elements SECIS tenen una estructura secundària característica en el mRNA que està altament conservada i conté seqüències consens que són indispensables per la incorporació de Sec. Estudis estructurals descriuen aquestes estructures consens (Figura 1).
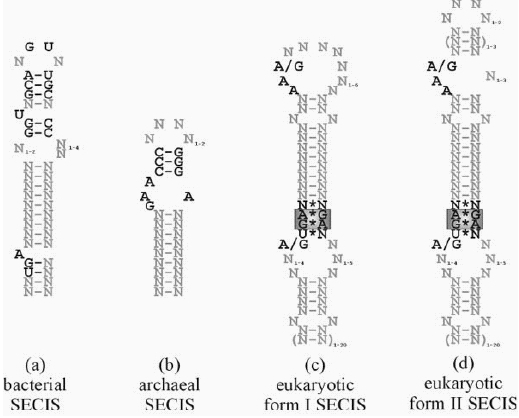 Figura 1.
Figura 1.torna a dalt
Selenoproteïna H
La selenoproteïna H (SelH) és una proteïna que en mamífers pesa aproximadament 14 kDa sense cap homologia amb cap proteïna funcionalment coneguda. En humans el gen que la codifica (ENSG00000211450) es troba al cromosoma 11 i ocupa aproximadament 1000 bases. Comparteix certes característiques amb la resta de selenoproteïnes, com el motiu CXXU. Aquest motiu també es troba en altres selenoproteïnes de mamífers com SelW, SelM, SelT i SelV. Recentment s'ha caracteritzat la funció de SelH i es creu que podria tractar-se d'una oxirreductasa nucleolar amb un patró molt inusual.
Mitjançant models in silico s'ha trobat homologia amb tiorreductases, ja que s'ha pogut identificar el mateix plegament en SelH. També per anàlisis de seqüències s'han identificat senyals de localització nuclear a la seqüència de SelH.
Pel que fa a l'expressió tissular, a través d'anàlisis d'expressió in silico s'ha trobat expressió moderada de mRNA de SelH en molts teixits i òrgans del cos. Això ha estat confirmat posteriorment per Northern Blot en teixits embrionaris i adults de ratolí. Anàlisis posteriors han demostrat que l'expressió de SelH es troba elevada en alguns tumors, especialment de tiroides, pulmons i fetge.
torna a dalt
Selenoproteïna I
La selenoproteïna I (SelI) està formada per 397 aminoàcids i conté una selenocisteïna a la posició 387 propera a la regió C terminal. L'mRNA de SelI va ser detectat en diferents teixits i tipus de cel·lulars.
Es coneix que la seva estructura gènica està constituïda per 10 exons i que el gen que codifica per aquesta proteïna es troba localitzat al cromosoma 2p23.3.
Recentment s'ha vist que SelI catalitza la biosíntesi de fosfatidiletanolamina a partir de CDP-etanolamina. Això juga un paper central en la formació i manteniment de les membranes vesiculars. Està involucrada en la formació de fosfatidiletanolamina a través de la via "Kennedy".
torna a dalt
Selenoproteïna L
La selenoproteïna L (SelL) conté dos residus de Sec altament conservats en un motiu UXXU. En eucariotes s'han trobat setze seqüències d'aquesta família: onze en peixos, dos en ascidis, dos en crustacis i una en mol·luscs. En procariotes s'ha trobat en dos microorganismes marins no identificats. Contràriament a la majoria de proteïnes en eucariotes, SelL està absent en mamífers, tot i que sí que s'han trobat homòlegs distants en Cys. S'ha identificat un únic element SECIS, també a la regió 3' UTR.
Construint un arbre filogenétic amb SelL, els seus homòlegs en Cys, i tiol oxidoreductases relacionades es va descobrir que SelL forma una família separada, diferent de les altres tiol oxidoreductases. Els homòlegs més propers a SelL són les proteïnes fmHP, que contenen un motiu CXXC en el lloc actiu predit. Les proteïnes fmHP s'han trobat en plantes, peixos i mamífers.
torna a dalt
Selenoproteïna N
La selenoproteïna N (SelN) està codificada pel gen SEPN1, el qual es troba al braç curt del cromosoma 1 a la posició 36.13. Mitjançant anticossos policlonals contra SEPN1 s'ha pogut determinar que el producte principal d'aquest gen és una proteïna de 70 KD que conté un únic residu de Sec. A més, amb experiments subcel·lulars s'ha pogut veure que es tracta d'una glicoproteïna localitzada a l'interior del reticle endoplasmàtic. L'expressió diferencial de SEPN1 depèn majoritàriament de regulacions post-transcripcionals.
Mutacions en aquest gen estan involucrades en desordres neuromusculars recessius, anomenats miopaties relacionades a SEPN1. De fet, SelN és la única proteïna involucrada en una malaltia genètica humana, però la seva funció encara no es coneix del tot.
SelN es detecta tempranament en precursors musculars i té un paper important durant l'embriogènesi, on actua des de la meitat de la gestació fins al període perinatal. Es creu que SelN pot ser important per les funcions musculars després del naixement, tot i que en teixits adults és activa a nivells molt més baixos.
SelN té similaritats estructurals amb altres selenoproteïnes, les quals estan involucrades en l'homeostasi redox. S'ha vist que SelN protegeix les cèl·lules humanes contra l'estrès oxidatiu, ja que una manca d'aquesta proteïna provoca un augment en l'activitat oxidativa de les cèl·lules. A més, SEPN1 juntament amb RyR1 (component principal del receptor intracel·lular de rianodina dels canals d'alliberament de calci) estan involucrats en els mateixos esdeveniments de la diferenciació cel·lular, i ambdós gens es necessiten per aconseguir fluxos adequats de calci dins l'embrió. Això té sentit ja que s'ha trobat una regió de SelN que té afinitat pel calci. S'ha vist que en absència de SEPN1 els receptors de rianodina tenen alterades les seves propietats bioquímiques i la seva sensibilitat per les condicions redox.
torna a dalt
Protists
Els protists (Protista) són un grup divers de microorganismes eucariotes. Històricament, els protists van ser considerats com un regne diferent, però aquest grup ja no és reconegut en la taxonomia actual. Els protists no presenten gaires característiques en comú a part d'una organització relativament senzilla: o bé són unicel·lulars o bé són multicel·lulars sense teixits especialitzats. Aquesta organització cel·lular simple diferencia els protists de la resta d'eucariotes, com ara els fongs, els animals i les plantes. De fet, el seu estil de vida és bastant similar al dels procariotes. La majoria d'organismes eucariotes són protists.
Els protists eren subdividits tradicionalment en diversos grups segons les seves semblances amb els regnes "superiors": els protozous unicel·lulars, semblants als animals; els protofitins, majoritàriament algues unicel·lulars semblants a les plantes; i els mixomicets i oomicets, semblants als fongs. Com que aquests grups sovint s'encavalquen, han estat substituits per classificacions basades en la filogènia. Tanmateix, encara resulten útils com a noms informals per descriure la morfologia i ecologia dels protists.
Els protists viuen en gairebé qualsevol medi que contingui aigua líquida. Molts protists, com ara les algues, són fotosintètics i productors primaris essencials dels ecosistemes, especialment a l'oceà, on formen part del plàncton. Altres protists, com ara els cinetoplàstids i els apicomplexos, són la causa d'una varietat de malalties humanes greus, com ara la malària i la malaltia de la son.
Els protists dels quals hem analitzat els genomes són els següents:
- Thalassiosira pseudonana
- Phytophtora sojae
- Phytophtora ramorum
- Theileria annulata
- Theileria parva
- Babesia bovis
- Entamoeba histolytica
- Entamoeba terrapinae
- Trypanosoma cruzi
- Monosiga brevicollis
- Giardia intestinalis
- Aureococcus anophagefferens
- Plasmodium berghei
- Plasmodium chabaudi
- Neospora caninum
- Toxoplasma gondii GT1
- Eimeria tenella
- Perkinsus marinus
- Dictyostelium purpureum
- Entamoeba dispar
- Entamoeba invadens
- Leishmania mexicana
- Leishmania braziliensis
- Emiliania huxleyi
torna a dalt