Thalassiosira pseudonana
 | Súper regne | Eukaryota |
|---|---|---|
| Fílum | Bacillariophyta | |
| Classe | Coscinodiscophyceae | |
| Família | Thalassiosiraceae | |
| Gènere | Thalassiosira | |
| Espècie | pseudonana |
Thalassiosira pseudonana és una diatomea cèntrica marina. Va ser el primer fitoplàncton eucariòtic marí en el que es va seqüenciar el seu genoma sencer.
Les diatomees són algues eucariòtiques unicel·lulars i fotosintètiques amb plastidis adquirits per endosimbiosi secundària. En l'endosimbiosi secundària, un eucariota no fotosintètic obté un cloroplast englobant un eucariota fotosintètic, probablement una alga vermella.
Les diatomees es troben en tots els oceans i aigües dolces del món (el gènere Thalassiosira en concret es troba en tots els oceans del món). Són la base de xarxes alimentàries curtes i eficients que mantenen piscifactories costeres.
El paper de les diatomees en l'ecologia i biogeoquímica del nostre planeta és crucial. Són responsables del 20% de la fixació del carboni terrestre a través de la fotosíntesi, convertint-se així en els organismes existents que més contribueixen al cicle global del carboni. Les diatomees, a més, dominen el cicle biogeoquímic del silici en el mar.
El tret més característic de les diatomees és la seva paret cel·lular de sílice anomenada frústula, que les ajuda a protegir-se de l'esclafament durant la predació. La frústula es composa de dos meitats de diferent mida i mostra nanoestructures específiques d'espècie tan detallades que han fet que les diatomees hagin estat utilitzades ja des de fa temps per testar la resolució dels microscopis òptics. En els últims anys s'ha prestat especial atenció a la biosíntesi d'aquestes nanoestructures com un paradigma per a la nanotecnologia de sílice del futur.
La llarga història (més de 180 milions d'anys) i dominància de les diatomees en els oceans es reflecteix per les seves contribucions als dipòsits de diatomita i a una fracció considerable de les reserves de petroli actuals.
T. pseudonana té un genoma d'una grandària de 29.453.142 nucleòtids.
Theileria parva
 | Súper regne | Eukaryota |
|---|---|---|
| Fílum | Apicomplexa | |
| Classe | Aconoidasida | |
| Ordre | Piroplasmida | |
| Família | Theileriidae | |
| Gènere | Theileria | |
| Espècie | parva |
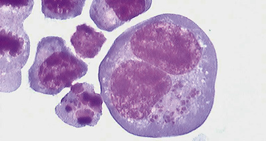
Theileria parva és un paràsit que es transmet per paparres i causa una malaltia del bestiar que es coneix com Febre de la Costa de l'Est (ECF) i es tracta d’un desordre limfoproliferatiu. T. parva pot infectar vaques, búfals Africans i búfals de la India.
La Febre de la Costa de l'Est, que mata aproximadament un milió de bestiar cada any a l'Àfrica Sub-Sahariana, resulta en pèrdues econòmiques que sobrepassen els 200 milions de dòlars anualment. La morbiditat i la mortalitat per ECF són atribuïdes a l'habilitat de l'esquizont (forma del paràsit) de transformar malignament la cèl·lula hoste, el limfòcit. El bestiar mor de malaltia limfoproliferativa de 3 a 4 setmanes després de la infecció.
La parasitosi augmenta exponencialment perquè l'esquizont es divideix sincrònicament amb la cèl·lula hoste i les cèl·lules infectades infiltren tots els teixits.
La transformació de les cèl·lules T per T.parva és el mecanisme predominant en la patogènesi de la ECF, però T.parva també transforma cèl·lules B. Quan el paràsit envaeix les cèl·lules T, aquestes pateixen una transformació limfoblastoide, es tornen independents de l'estimulació del receptor per antígens i deixen de necessitar factors de creixement exògens per a proliferar. Les cèl·lules T transformades per T.parva mostren moltes característiques de les cèl·lules tumorals, proliferant incontroladament.
Els organismes Theileria pertanyen al fílum Apicomplexa, que es pensa que es va originar fa 930 milions d'anys.
El genoma de T.parva conté 8.347.606 nucleòtids.
Trypanosoma cruzi
 | Súper regne | Eukaryota |
|---|---|---|
| Fílum | Sarcomastigophora | |
| Classe | Euglenozoa | |
| Ordre | Kinetoplastida | |
| Família | Trypanosomatidae | |
| Gènere | Trypanosoma | |
| Espècie | cruzi |
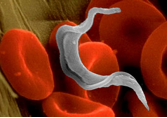
T.cruzi és un paràsit intracel·lular d'hostes vertebrats. Es troba exclusivament a Amèrica i produeix una zoonosi anomenada malaltia de Chagas.
Els tripanosomàtids tenen estructures cel·lulars úniques entre els protozoaris. Per exemple, una de les característiques que destaquen d'aquests organismes és el conjunt de microtúbuls subpel·liculars, els quals es troben adossats a la membrana citoplasmàtica (pel costat intern), conformant un citoesquelet perifèric de gran rigidesa. Els microtúbuls subpel·liculars es distribueixen en tota la membrana citoplasmàtica, excepte en l'àrea d'on surt el flagel. Aquesta regió carent de microtúbuls és de gran importància per a la cèl·lula, ja que és l'únic lloc on es realitza endocitosi o exocitosi de molècules.
Una altra estructura especialitzada de T.cruzi i altres tripanosomàtids és el cinetoplast, que és una xarxa d'ADN extranuclear localitzada en un punt específic del mitocondri. Aquesta xarxa d'ADN representa una proporció important de l'ADN total cel·lular, ja que, depenent de l'espècie, pot contenir del 10 al 20% de l'ADN total de la cèl·lula.
El gènere Trypanosoma es caracteritza per utilitzar dos hostes, un vertebrat i un altre invertebrat, per a completar el seu cicle de vida. Existeixen 4 formes morfològiques principals dels tripanosomes: tripomastigot, epimastigot, promastigot i amastigot. Les diferents formes es distingeixen entre si per la posició del cinetoplast en relació amb el nucli i per la presència o absència d'una membrana ondulant.
La infecció per T.cruzi és transmesa per xinxes a més de 100 espècies diferents d'animals inclosos els humans. Els xinxes s'infecten en picar un animal infectat, ingerint així al paràsit. La infecció del mamífer s'inicia quan un insecte infectat defeca alliberant tripanosomes en la seva femta i orina. Els tripanosomes, incapaços de travessar la pell intacta, entren a l'organisme a través d'excoriacions de la pell (lloc de la mossegada), o a través de les mucoses, envaint immediatament les cèl·lules hostes.
La infecció es localitza únicament a Amèrica, des del sud d'Estats Units fins a Argentina i Xile. La infecció per T.cruzi té una fase aguda inicial amb una duració de diverses setmanes i una fase crònica que persisteix durant la vida de l'hoste. Durant la fase inicial de la infecció el paràsit es multiplica ràpidament ja que no hi ha ni reacció inflamatòria al voltant de les cèl·lules parasitades, ni una resposta immune específica. Els paràsits es disseminen a través de la circulació. T.cruzi està dividit en dos subgrups genètics divergents, els llinatges I i II, que es creu que van divergir fa entre 88 i 37 milions d'anys. Aquests dos llinatges inclouen tot tipus de soques aïllades d'humans, insectes que actuen com a vectors i mamífers salvatges.
T.cruzi presenta un genoma de 89.612.356 nucleòtids.
Theileria annulata
 | Súper regne | Eukaryota |
|---|---|---|
| Fílum | Apicomplexa | |
| Classe | Aconoidasida | |
| Ordre | Piroplasmida | |
| Família | Theileriidae | |
| Gènere | Theileria | |
| Espècie | annulata |
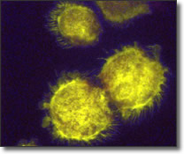
Theileria annulata és un paràsit intracel·lular que causa una malaltia anomenada Theileriosi Tropical en el bestiar de països tropicals i subtropicals, on 250 milions de bestiar estan en risc d'infecció.
La infecció provoca una malaltia limfoproliferativa ja que el paràsit trastorna la senyalització de les cèl·lules (leucòcits i eritròcits) que envaeix. La Theileriosi s'acompanya típicament d'anèmia severa.
La malaltia es transmet per paparres i té una alta morbiditat i mortalitat. T.annulata pot infectar vaques, búfals de la India i camells.
T.annulata es distribueix pel Nord d’Àfrica, Sud d’Europa, i per Àsia.
Phytophthora sojae
 | Súper regne | Eukaryota |
|---|---|---|
| Fílum | Oomycota | |
| Classe | Oomycetes | |
| Ordre | Peronosporales (Phythiales) | |
| Família | Pythiaceae | |
| Gènere | Phytophthora | |
| Espècie | sojae |
Phytophthora és un gènere del que se’n coneixen 59 espècies i que, a través d'evolució convergent, té similaritats amb els fongs. És un gènere que ataca centenars d'espècies diferents de plantes, incloent-hi molts conreus, provocant danys amb costos de desenes de bilions de dòlars l’any.
P.sojae és un oomicet que va ser identificat a Ohio i a Carolina del Nord l’any 1954. Des de llavors, el dany per aquest organisme s’ha observat a Argentina, Austràlia, Brasil, Canadà, Hongria, Itàlia, Japó i a molts altres indrets.
Les temperatures càlides i els sòls saturats d’aigua afavoreixen la infecció per P.sojae i el desenvolupament d’una malaltia que es caracteritza per pèrdues de resistència i reduccions en el rendiment de cultius de soja, portant a la mort en el cas dels cultius més susceptibles.
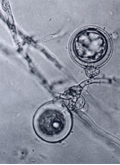
Phytophthora sojae sobreviu com oospores al terra i en restes de plantes. Les oospores poden sobreviure uns quants anys en estat latent i poden resistir la congelació i períodes llargs de fred. A la primavera, les oospores germinen sempre que la temperatura sigui l’adequada per formar esporangis. Els esporangis o germinen directament o germinen indirectament acumulant-se fins que el sòl és inundat d’aigua, moment en el que s’alliberen les zoospores. Les zoospores són atretes cap a les llavors i arrels per exsudats d’isoflavonoides. Un cop arriben a l'arrel, les zoospores perden el seu flagel, produeixen una paret cel·lular (encist), germinen, i subseqüentment infecten els teixits de l'arrel formant-se oogònies i oospores a l’arrel infectada.
Per pal·liar els efectes que P.sojae ocasiona en els cultius de soja es poden utilitzar cultius resistents específics de raça.
P.sojae té un genoma mitocondrial de 42.977 nucleòtids.
Phytophthora ramorum
 | Súper regne | Eukaryota |
|---|---|---|
| Fílum | Oomycota | |
| Classe | Oomycetes | |
| Ordre | Peronosporales (Phythiales) | |
| Família | Pythiaceae | |
| Gènere | Phytophthora | |
| Espècie | ramorum |
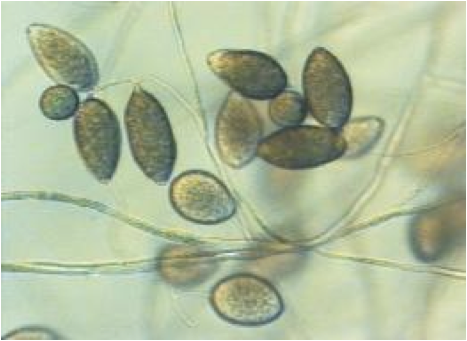
És un fong patogen que infecta arbres, arbusts i altres plantes.
Des dels anys 90 ha sigut el causant de la mort de milions d’arbres a la costa americana. La majoria d’arbres infectats eren roures, i per això la infecció amb aquest fong se la coneix com “la mort sobtada dels roures”.
Aquest organisme es transmet produint dos tipus d’espores resistents que permeten la supervivència i expansió del fong. Les espores que arriben a les fulles mullades, germinen i infecten la planta. Els òrgans de reproducció asexual comprenen els esporangis, zoospores i clamidospores. Aquestes últimes són importants per sobreviure a les condicions desfavorables. S'ha observat que les fulles més joves són especialment susceptibles a aquestes espores.
La major quantitat d’hostes de P.ramorum es troba als Estats Units (nord-est), on s'han registrat 29 espècies de plantes infectades amb unes 31 espècies associades. A Europa també s'han registrat infeccions en diverses plantes d’Alemanya, Bèlgica, França, Polònia, Espanya, Itàlia, Suïssa i Anglaterra.
Els símptomes són diferents segons el tipus d’hoste. En algunes plantes, el teixit foliar es torna totalment necròtic; mentre que en d’altres, només es presenten petites lesions en les fulles més baixes.
EL DNA mitocondrial de P.ramorum mesura aproximadament 39314 nucleòtids i té 71 gens.
Monosiga brevicollis
 | Súper regne | Eukaryota |
|---|---|---|
| Fílum | Sarcomastigophora | |
| Classe | Zoomastigophora | |
| Ordre | Choanoflagellida | |
| Família | Codonosigidae | |
| Gènere | Monosiga | |
| Espècie | brevicollis |
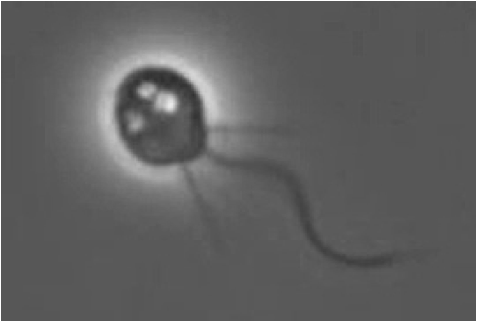
És un protista aquàtic unicel·lular, que pertany a l'ordre dels coanoflagelats. Els microorganismes d'aquest grup es caracteritzen per la presència d'un únic flagel rodejat per un anell amb de 30 a 40 microvellositats (microvilli). El moviment del flagel fa una corrent d'aigua que els permet moure's i alimentar-se de bacteris i detritus que resten atrapats en aquesta columna d'aigua.
Els coanoflagelats es poden trobar com organismes unicel·lulars o formant colònies petites. S'ha determinat que la seva abundància és relativa a la presència d'altres organismes com els nanoplàncton dins de les seves comunitats. Aquests organismes compleixen un rol primordial en la cadena d'alimentació microbiana i en el cicle del carboni.
Els científics han apuntat des de fa temps que els coanoflagelats podrien ser exemples moderns de com eren els ancestres dels animals multicel·lulars (metazous). L’evidència que dóna suport a aquesta hipòtesi, és que els coanoflagelats són gairebé idèntics a les cèl·lules de les esponges, anomenades coanòcits, que també realitzen la funció de recol·lecció d'aliment.
S'han fet diferents estudis genòmics per a comprovar el grau de parentiu entre els coanoflagelats i els animals superiors. Els científics han comparat gens de M.brevicollis amb quatre gens animals que expressen proteïnes que estan altament conservats al llarg del regne animal (factor d’elongació 2, alfa tubulina i actina). Els resultats d'aquestes comparacions han confirmat la relació entre aquests protozous i els metazous.
El DNA genòmic de M.brevicollis mesura 38648938 nucleòtids i presenta 9174 gens.
Giardia intestinalis
 | Súper regne | Eukaryota |
|---|---|---|
| Fílum | Sarcomastigophora | |
| Classe | Eopharyngia | |
| Ordre | Diplomonadida | |
| Família | Hexamitidae | |
| Gènere | Giardia | |
| Espècie | intestinalis |
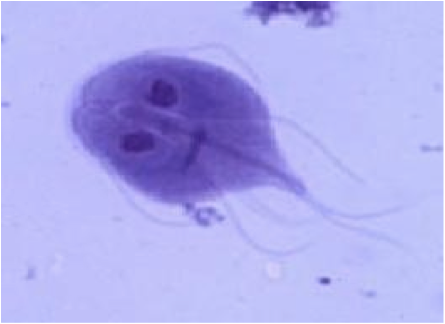
És un protozou flagel·lat intestinal, tres vegades més freqüent en nens que en adults. Aquest paràsit és el principal responsable de les parasitosis infantils.
Pertany al subfílum mastigophora, el que indica que són protozous que tenen flagels i membranes ondulants. Presenten quatre parells de flagels que neixen d'unes estructures anomenades blefaroblasts; aquestes estructures es designen en relació a la seva posició (anterior, posterior, neutral i cabal) i faciliten el diagnòstic del paràsit.
La Giardia intestinalis o Giardia lamblia, presenta dues formes morfològiques en el seu cicle evolutiu: el trofozoït o la forma mòbil i el quist, forma més petita que permet la supervivència de l’organisme en les condicions ambientals més adverses.
La forma mòbil té simetria bilateral i és piriforme, amb el seu extrem anterior ample i l'extrem posterior molt prim. A la part anterior presenta una estructura anomenada disc suctor, que li permet adherir-se a l'epiteli intestinal; a més, a prop del disc suctor hi ha dos nuclis idèntics ovalats amb una enorme massa de cromatina central. L'adhesió a l'epiteli intestinal també té lloc gràcies a una estructura anomenada axolema o axostil, que fa d'esquelet. A sobre de l’axostil es troben els cossos parabasals.
El quist és una estructura ovalada i petita que mesura 6-7 micres. Presenta una paret molt gruixuda anomenada paret quística, que és fonamental per a la resistència a les diferents condicions del medi ambient; aquesta paret està formada per una capa filamentosa externa i una capa membranosa interna.
Cicle Biològic:
Els quists són les formes resistents i responsables de la transmissió de la giardiasi. Aquests són transmesos a l'hoste mitjançant l'aigua o aliments contaminats.
S'estima que d’uns 10 a 100 quists són suficients com a dosi infectant. Després de la ingestió del quist, el contacte del quist amb els sucs gàstrics indueix l’activació d’aquest.
En resposta a pH alcalí, les proteases de l’intestí i senyalitzacions pròpies del paràsit, emergeix una cèl·lula que dóna origen a trofozoïts tetranucleats, els quals es divideixen originant dos trofozoïts binucleats. Aquests es poden trobar de forma lliure al duodè o units a la mucosa duodenal gràcies al disc suctor.
L'enquistació té lloc a mesura que el paràsit és arrossegat pel trànsit intestinal cap al colon, i aquests quists sortiran cap a l’exterior amb les femtes. En alguns casos també poden sortir com trofozoïts quan no hi ha hagut temps de transformar-se en quist, però en sortir com trofozoït el paràsit patirà una desintegració ja que no serà resistent a les condicions de l’exterior.
L’adhesió del paràsit a la mucosa intestinal és el mecanisme de dany de la infecció amb giardia, ja que en adherir-se fortament a l’epiteli genera una absorció deficient dels nutrients. A més, s'ha observat la secreció de moc producte de la irritació que produeix la presència dels trofozoïts a la paret. La generació d’aquest moc, contribueix també a l'obstrucció de l'absorció i a una major reacció inflamatòria.
El genoma de Giardia intestinalis, mesura aproximadament 111922 nucleòtids i està format per 6583 gens.
Entamoeba terrapinae
| Súper regne | Eukaryota |
|---|---|
| Fílum | Sarcomastigophora |
| Classe | Lobosa |
| Ordre | Schizopyrenida |
| Família | Entamoebidae |
| Gènere | Entamoeba |
| Espècie | terrapinae |
És un protozou comensal d’alguns tipus de tortugues. Aquest protozou pertany al gènere Entamoeba, on es troben la majoria de les amebes endoparàsites. Totes les espècies d’aquest gènere (a excepció de la E. gingivalis) presenten una fase quística, on es produeixen varies divisions del nucli, amb el mateix nombre de divisions del citoplasma.
Aquests paràsits presenten un cicle directe, en el que colonitzen el tracte gastrointestinal de les tortugues provocant diarrea, deshidratació, etc. Les lesions més freqüents observades són gastritis, ulceració i necrosi de la mucosa gastrointestinal.
La infecció s'origina per la ingestió de quists infectants que es troben en les femtes d’altres rèptils infectats.
Entamoeba histolytica
 | Súper regne | Eukaryota |
|---|---|---|
| Fílum | Sarcomastigophora | |
| Classe | Lobosa | |
| Ordre | Schizopyrenida | |
| Família | Entamoebidae | |
| Gènere | Entamoeba | |
| Espècie | histolytica |
És un protozou comensal que pertany al subfílum sarcodina, que inclou els protozous que emeten diferents tipus de pseudòpodes com a medi de locomoció.
Aquest protozou és un paràsit obligat de l’intestí gros humà, que en alguns casos envaeix la mucosa intestinal i pot disseminar-se per via hepàtica. És l'agent responsable de l’amebiasi, que és una afecció cosmopolita.
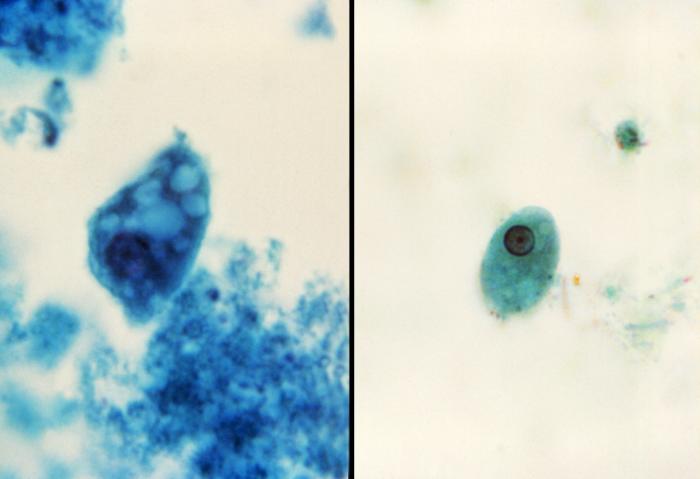
E.histolytica presenta dues formes al llarg del seu desenvolupament: el trofozoït (fase invasiva) i el quist (fase infectant).
El trofozoït és mòbil i completament pleomòrfic, perquè el seu aspecte i mobilitat estan influenciats pels canvis de pH, potencial redox i osmolaritat. Es multiplica per fissió binària. El seu hàbitat es troba a la paret del còlon humà. La seva mida oscil·la entre els 10 i 60 μm; les formes més petites corresponen a les no invasives, i són les que es troben en els casos asimptomàtics. Per altra banda, les de major mida corresponen a les formes invasives. El trofozoït presenta una membrana citoplasmàtica dividida en dos: l'ectoplasma (d'on surten prolongacions que corresponen als pseudòpodes) i l'endoplasma. La nutrició es fa per fagocitosi de teixits dissolts i alguns hematies.
Si les condicions del medi no són favorables, el trofozoït comença a canviar de forma i deixa d'emetre pseudòpodes, l'ectoplasma i endoplasma ja no es diferencien, i es forma una paret externa molt gruixuda (paret quística). Tot aquest canvi porta a la formació del quist. El quist o element infectant és rodó i de 10 -25 μm de mida. Presenta una paret llisa i és resistent al suc gàstric i factors externs. Segons la fase de maduració té de 1 a 4 nuclis. Els quists joves tenen de 1 a 2 nuclis, i quan aquests maduren tenen 4 nuclis.
La infecció per E.histolytica es dóna tan en climes freds com en zones tropicals, però en general, les àrees amb clima temperat o calent són les de major endèmia. Aquestes infeccions són més freqüents en homes que en dones; a més, la prevalença està influenciada pel nivell higiènic-sanitari de la població i medi ambient.
Pel seu mecanisme de transmissió, la amebiasi pertany al grup de les protozoosis transmeses per fecalisme, perquè les formes infectants s'ingereixen amb aliments o begudes contaminats amb materials fecals de persones parasitades amb el protozou amb o sense simptomatologia (persones portadores).
Cicle Biològic:
Els quists són transmesos mitjançant les femtes. La infecció per E.histolytica té lloc per la ingestió de quists madurs que es troben a l'aigua o a aliments contaminats amb matèria fecal.
Quan ja s'ha ingerit el quist madur, aquest arriba a l'intestí, on s'inicia el procés de desenquistament induït pel contacte amb els sucs gàstrics: els nuclis es multipliquen fins a 8 i s'alliberen petites formes trofozoïques anomenades amèbules metaquístiques, que es desenvolupen a trofozoïts madurs. Aquests últims migren a l'intestí gruixut.
Els trofozoïts es multipliquen per fissió binària i formen quists, que seran expulsats amb les femtes. Gràcies a la protecció de la paret, els quists poden sobreviure dies o setmanes en el medi exterior i són els responsables de la transmissió de la malaltia.
En alguns casos, els trofozoïts es mantenen al lumen intestinal (infecció no invasiva), en aquest cas, els individus són portadors asimptomàtics que expulsen quists amb les femtes. En altres pacients, els trofozoïts envaeixen la mucosa intestinal (malaltia intestinal) o mitjançant el flux sanguini poden arribar al fetge, cervell o pulmons (malaltia extraintestinal).
El DNA genòmic de E.histolytica mesura 20772429 nucleòtids i té 8342 gens.
Babesia bovis
 | Súper regne | Eukaryota |
|---|---|---|
| Fílum | Microspora | |
| Classe | Microsporea | |
| Ordre | Piroplasmida | |
| Família | Babesiidae | |
| Gènere | Babesia | |
| Espècie | bovis |
És un protozou intraeritrocitari que pertany al fílum apicomplexa, subclasse piroplasmida i són coneguts com piroplasms per la seva forma de “pera” en els seus estadis de merozoïts.
La infecció amb aquests paràsits provoca Babesia, una malaltia semblant a la malària humana. Els paràsits infecten bovins, ovins, gossos i ocasionalment humans. La infecció humana va ser reportada a Europa i Nord Amèrica i se la coneixia com la “febre de la paparra”. La babesiosi humana és una zoonosi, adquirida per la picada de paparra infectada.
Cicle biològic:
Tots els membres del gènere Babesia, presenten dos cicles de vida: La fase sexual, quan el paràsit es troba dins de l’hoste invertebrat (paparra) i la fase asexual, quan es troba dins de l'hoste vertebrat.
Els esporozoïts s'introdueixen al flux sanguini de l’animal mitjançant la picada d’una paparra infectada. Els esporozoïts entren en els eritròcits, on per fissió binària es transformen en trofozoïts. Per gemmació (reproducció asexual) s’alliberen a la sang merozoïts, i la majoria infecten nous eritròcits. A més, alguns merozoïts es diferencien en gàmetes masculins i femenins.
Quan una paparra sana pica a un animal infectat, els gàmetes que es troben a la sang, arriben fins a l'intestí de la paparra, on es fusionen formant el zigot. Els zigots a la vegada es converteixen en oocinets mòbils que penetren les glàndules salivals del vector. S'inicia llavors un cicle d’esporogònia que dóna com a resultat nous esporozoïts, que poden ser injectats a un altre animal per la picada de la paparra.