|
||||||
| Abstract |
Introduction | Materials and Methods |
Results | Discussion | Conclusions | References |
|
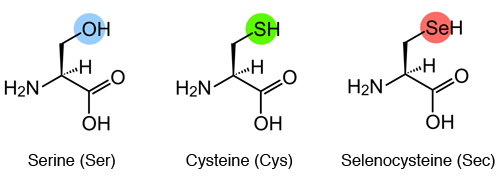
IntroductionSelenium is an essential micronutrient for the life of many organisms. In humans, both excesses and deficiences of this nutrient can lead to diseases. When selenium deficiency appears, it leads to Keshan disease which affects the cardiac system, whereas selenium excess can lead to poisoning. In nature, selenium is not found elementally. To prevent its toxicity it binds to a selenocysteine residue, present in selenoproteins. [1] SelenocysteineSelenocysteine is the amino acid number 21, it is encoded by the TGA codon and is represented by two abbreviations: Sec and U. Structurally, selenocysteine and cysteine are very similar but selenocysteine has a selenium atom (SeH group) replacing the sulfur atom (Sulfhydryl or SH group) of cysteine. The functional similarity between selenocysteine and cysteine due to evolutionary processes allows the finding of orthologue protein sequences containing selenocysteine replaced by cysteine. [2]
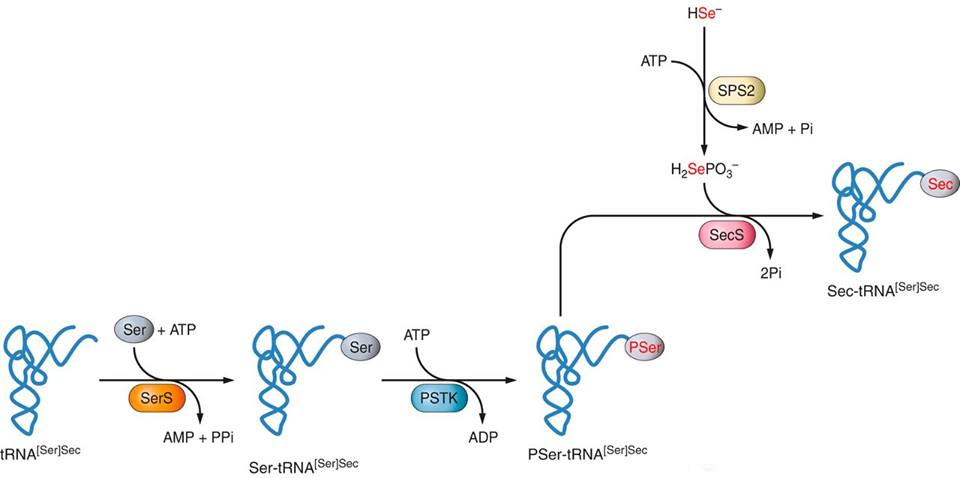
Selenproteins and their phylogenySelenoproteins are, mostly, enzymes that present a selenocysteine in their active site. There is a large number and variety of selenoproteins distributed in the three domains of life: bacteria, archaea and eukaryotes, but not in plants. Furthermore, it was observed that there are animals that do not have these selenoproteins present. 45 families of selenoproteins have been described so far. In fish there are up to 41 of these families and in mammals up to 28 although one mammal can have up to 25 mammalian selenoproteins in its genome. [3] Broadly speaking, selenoproteins act as redox enzymes and are believed to have antioxidant, antitumoral and immune function as well as selenium chelators to reduce toxicity. However, the function of some selenoprotein families is unknown and the distribution of selenoproteins along the evolutionary line is incomplete. [4] Synthesis of selenoproteinsThe machinery responsible for regulating the synthesis of selenocysteines and selenoproteins comprises the following enzymes: SPS1, SPS2, SecS, Pstk, eEFsec and SBP2. [5] Synthesis of selenocysteineThe only form of selenium capable of being part of selenocysteine is selenophosphate (H2PO4SeH) obtained through the modification carried out by Selenophosphate synthetase enzyme (SPS2) of dietary selenium intake. The structure responsible for incorporating the selenocysteine to the amino acid chain in formation during protein translation is selenocisteinil-tRNA (tRNA + selenocysteine), also called tRNA[Ser]Sec. Selenocysteine is the only amino acid that is synthesized directly at the tRNA and it is like follows:
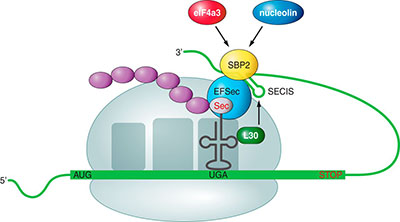
Incorporation of selenocysteine during translationUsually, the UGA codon (corresponding the TGA codon in DNA) is a STOP codon. But in the coding sequences for a selenoproteins UGA is not played as a STOP by the transcriptional machinery so the protein incorporates the selenocysteine and continues until the next STOP codon. [5][6] In eukaryotes, the mechanism by which this codon is interpreted differently is mediated by the presence in the extreme 3 'UTR of a three-dimensional structure in a RNA loop called SECIS (SElenoCystein Insertion Sequence). This sequence allows the incorporation of selenocysteine in a chain of amino acids in formation through the following mechanism:
Selenoprotein familiesDIIodothyironine deiodinases (DI) are a group of homodimeric selenoproteins that are related to the metabolism of thyroid hormone. They catalyze the conversion of pro-hormone T4 into active isomers of triiodothyronine (T3), inactive reverse triiodothyronine (rT3) and diiodotironina (T2). This biotransformation of the thyroid hormone occurs in almost all tissues and is catalyzed by three different isotypes of desiodinases: DI1 (liver and kidney), DI2 (heart, skeletal muscle, central nervous system, adipose tissue, thyroid and pituitary gland) and DI3 (fetal tissue and placenta). The deiodinases DI1 and DI2 are used to enable or to remove iodine residue from the outer ring (ORD) hormone. And DI3 mainly catalyzes the deactivation or the addition of a iodine residue to the inner ring (IRD) of the pro-hormone T4 and rT3 to T3 and T2, respectively. [7] GPxGlutathione peroxidase enzymes constitute a family of eight mammalian selenoproteins that are important in the cell cycle system of antioxidant defense and have a high conservation. The main function of glutathione peroxidases is to protect the body from oxidative and degrading effect of peroxides and hydroperoxides formed endogenously. This protein has a selenocysteine in the catalytic site, and regulates the disposal of selenium enzyme glutathione peroxidase. [8] Each GPX has a specific function and location:
MsrA i SelRMetionin sulfoxide reductase peptides, MsrA and MsrB (SelR), do not share homology either in structure nor in sequence, therefore, both have a different evolutionary origin. These peptides have a restorative function of proteins that have been inactivated by methionine oxidation. They catalyze the reduction of methionine-sulfoxide into methionine. In fact, MsrA is the only protein in mammals capable of carrying out this function (reducing metionenine-S-sulfoxide). In mammalian, MsrA contains cysteine and MsrB a selenocysteine. MrsA also contains a selenocysteine in prokaryotes. MsrB has three isoforms: SelR1, SelR2 and SelR3, among which SelR3 and SelR2 have higher homology. [9] SelHSelenoprotein H is an oxidoreductase thioredoxin-like protein with a nuclear CXXU conserved motif (a cysteine separated by two random residues from a selenocysteine). This motif suggests a redox role of SelH and also suggests that this role depends on the trace element selenium. Not known homology with other characterized selenoproteins. This structure is also present in several mammalian selenoproteins (SelW, SelT, SelM). [10] Sel ISelenoprotein I (Sel I) is a protein that contains a selenocysteine near the C-terminal region. It is known it has a CDP-alcohol phosphatidyltransferase domain very conserved that is typically found in the choline-phosphotransferase 1 (CHPT1) and choline/ethanolamine phosphotransferase (CEPT1). CHPT1 catalyzes the transfer of choline into diacylglycerol from CDP-choline amd CEPT catalyzes a similar reaction but accepts both choline and ethanolamine. Sel I catalyzes the biosynthesis of phosphatidylethanolamine from CDP-ethanolamine. This selenoproteins has 7 transmembrane predicted domains (similar to CHPT1 and CEPT1). An important sequence of the protein is between the first and second transmembrane domains, where three aspartic acid residues are located and which are critical for its function. [3] Sel KSelenoprotein K (SelK) is a transmembrane protein located in the endoplasmic reticulum with a high expression in certain tissues such as the heart, spleen, areas of the brain and some immune cells. It is found in many species of higher eukaryotes. Many studies suggest that SelK is involved in the regulation of calcium and the protein degradation pathway associated with the endoplasmic reticulum. This pathway and protein degradation system is responsible for eliminating proteins in the cytoplasm if these are not properly folded in the ER. It is believed that SelK has a well-defined structure because it has an SH3 domain and it is rich in glycine, proline and charged residues. It is currently studied whether its function is related to oxidative stress response in the endoplasmic reticulum. [11] Sel MSelenoprotein M (SelM) is a protein found in the endoplasmic reticulum. It has two regions of interest: a redox CXXU motif and a thioredoxin-like (TRX) folding. It plays an important role in the regulation of calcium and protection against oxidative stress. It is expressed mainly in the brain. In many vertebrate species this selenoprotein is lost.Terminal residues appear to have a KDAL retention sign in the endoplasmic reticulum as it is perinuclear. SelM is a distant homologue of Sel15. It is located in the ER / Golgi where it is associated with UDP-glucose (Glycosyltransferase glycoprotein), a protein located in the ER involved in the control of protein folding quality; controlling whether disulfide bonds are well formed.[5] Sel NSelenoprotein N (SelN) is a glycoprotein of the endoplasmic reticulum with an expression pattern of early development. It plays a key role in muscle function after birth; however, during adulthood it is expressed in very low concentrations. This selenoptoreïna consists of two isoforms are expressed in skeletal muscle, brain, lung and placenta. Isoform II is also expressed in the heart and stomach. Defects in gene expression encoding SelN (SEPN1) result in congenital muscular dystrophy with spinal stiffness (RSMD1). [12] Sel OSelenoprotein O (Sel O) is a protein that is widely distributed: it has homologies in animals, bacteria, yeast and plants. Sel O adopts a three-dimensional folding similar to the kinases folding, but without preserving the aspartate residue in the catalytic domain Arg-His-Asp. Possibly, this kinase domain gives the protein an important role in the cell signaling network. In yeast and bacteria, it has a role in the response to oxidative stress. There has also been an important role in the ABC transport and the regulation of metabolism. Only SelO homologues in vertebrates have selenocysteine, which are located in the penultimate position of the C-terminal end. Like SelM, the apical loop of the SECIS element has two cytosines instead of two adenines (observed in mammals). [13] Sel PSelenoprotein P (SelP) is a selenium-rich glycoprotein with multiple selenocysteine residues - 10 in humans - located in the N-terminal domain (proenzyme) and C-terminal (transport domain). It is the most common plasma selenoprotein. It is essential for the homeostasis of selenium in the body and promotes its retention in the body and affects selenium transport from the liver to extra-hepatic tissues. The SelP may also be involved in regulating glucose metabolism by regulating pancreatic insulin secretion. It also has an antioxidant effect due to the capability of joining heparin and heavy metals that present in the organism, such as cadmium and mercury. The C-terminal domain has 9 selenocysteines and the N-terminal domain has only 1. SelP has been associated with Alzheimer’s disease because it has been observed SelP colocalization within amyloid plaques. [14] Sel SSelenoprotein S (SelS) may have or have not SECIS elements depending on RNA splicing during its transcription. The expression of SelS is associated with cardiovascular disease and diabetes and it is regulated by the cellular response to glucose and insulin. It also has been associated with the regulation of the inflammatory response and therefore regulated by inflammatory citokines. In the membrane of the endoplasmic reticulum, it is involved in the retrotranslocation of misfolded proteins to the cytoplasm, where it will be degraded. It has to do with the response to oxidative stress. [15] Sel TSelenoprotein T (SelT) is a small selenoprotein with a putative redox N-terminal motif. Because of the presence of a hydrophobic domain it is suspected that its location is in the plasma membrane. [16]
Sel USelenoprotein U (SelU) is a selenoprotein found in fish and many other marine organisms, birds and algae; his homologue in cysteine is present in mammals, arthropods, worms, amphibians and terrestrial plants. In mammals, there have been predicted three different isoforms: SelU1, and SelU2 SelU3. Currently the function is unknown. SelU1 is expressed in tissues such as bone, brain, liver and kidney. [17] Sel WSelenoprotein W (SelW) is the smallest mammalian selenoprotein. It is expressed in different eukaryotes and bacteria, and its distribution is not limited to a single organ in mammals. We have seen that it could play a role in the regulation of mutagenesis, gastrointestinal tract and immune system. [18] TRThioredoxin reductases (TR) are proteins with sequence similar to those in the glutathione reductase. They have a conserved redox catalytic domain: Cys-Val-Asn-Val-Gly-Cys-. Its function is to reduce tiroreduccines. Mammals have three TR isoenzymes; TR1 is located in the cytoplasm and TR2 and TR3 have mitochondrial localization.[19] Balaenoptera acutorostrata
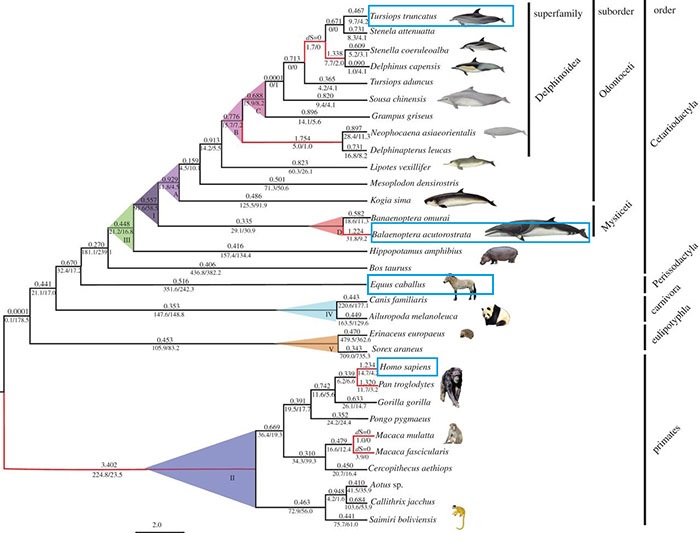
Balenoptera acutorostrata, commonly called common minke whale, is a large mammal that lives in big oceans and seas around the world. It is the smallest species of baleen whales and it can measure up to 10.2 meters long and weigh 10.000kg. Although they are not considered coastal, these whales are not more than 169km away from the coast and sometimes enter estuaries, bays, fjords and lakes. Its distribution is extensive and varied throughout the different oceans, but it prefers cold Polar Regions than tropical and temperate regions. Like most of the whales, the minke whale feeds mainly on krill and small fish but their diet can be omnivorous, especially those which inhabit regions further north. Usually they travel alone or in small groups (2-4), but are known to congregate in areas rich in krill. Gestation lasts 10 or 11 months and famales have only one baby per pregnancy. The cubs weigh 450kg at birth and feed on breast milk for 5 months. They do not reach sexual maturity until the age of 6.
Choose genome reference
|