
Nomascus leucogenys
Biologia
És una espècie de primat de la familia Hylobatidae.
Llinatge segons NCBI: cellular organisms; Eukaryota; Opisthokonta; Metazoa; Eumetazoa; Bilateria; Coelomata; Deuterostomia; Chordata; Craniata; Vertebrata; Gnathostomata; Teleostomi; Euteleostomi; Sarcopterygii; Tetrapoda; Amniota; Mammalia; Theria; Eutheria; Euarchontoglires; Primates; Haplorrhini; Simiiformes; Catarrhini; Hominoidea; Hylobatidae; Nomascus.
Els Nomascus leucogenys no presenten diferències de mida entre mascles i femelles, tots dos pesen al voltant de 5.7kg, així mateix, medeixen entre 45 i 63 cm. En canvi, els Nomascus leucogenys són dimòrfics en color de pell. Tots els infants neixen de color beige, als dos anys canvien de beige a negre i desenvolupen taques blanques a les galtes. Quan arriben a la maduresa sexual, els mascles conserven el color negre amb les galtes blanques, mentre que les femelles tornen al seu color beige original. Com les altres espècies de gibons, els Nomascus leucogenys no tenen cua, però tenen una extremitats excepcionalment llargues, el seu cos està adaptat per a viure als arbres.

Distribució, habitat i ecologia
Viuen en els boscos de Vietnam, Laos, i la provincia xina de Yunnan, en zones de clima subtropical, amb hiverns curts i sense gelades. Es considera una espècie en perill d'extinció, degut a que hi ha una alta taxa de caça i pèrdua del seu habitat. Els Nomascus leucogenys són principalment arborícoles i frugívors, però encara hi ha poques dades sobre la seva ecologia. Es va fer un estudi en que s'analitzava el contingut de l'estomac de sis N. leucogenys, i es va observar que del 90 al 100% dels residus eren fruites associades a alguns insectes i fulles d'arbres. També s'ha vist que en la temporada de pluges, quan hi han més fruites disponibles, els N. leucogenys viatgen menys; mentre que en l'estació seca, els N. leucogenys viatgen distàncies més llargues.
Els Nomascus leucogenys són monògams, arriben a la maduresa sexual al voltant dels 7 anys d'edat. En aquest moment, les femelles comencen a tenir un cicle menstrual que dura 28 dies. Es reprodueixen durant tot l'any i quan es produeix la fecundació, la femella té un periòde de gestació de 7 mesos. Quan el nadó neix s'aferra a la mare durant gairebé 2 anys, moment en què és deslletat.
Seqüenciació del genoma
El genoma del Nomascus leucogenys ha estat seqüenciat per la Universitat de Washington en col·laboració amb Baylor University College of Medicine Human Genome Sequencing Center (BCM-HGSC). La seqüenciació s'ha realitzat a partir del genoma d'un Nomascus leucogenys salvatge anomenat Asia, que servirà com a referència del genoma dels gibons.
A excepció dels ximpanzés, goril·les i orangutans, els gibons són filogènicament els parents més propers dels humans, en comparació amb altres primats hominoides. El genoma del gibó és l'únic que té nombrosos reordenaments intercromosomals. El projecte de seqüenciació del genoma del gibó identificarà aquests reordenaments mitjançant seqüenciació de BAC-end i ha de proporcionar importants coneixements sobre els processos d'evolució i especiació. Els gibons han estat recentment reclassificats en gèneres pel seu nombre diploide de cromosomes, el gènere Nomascus té 52 cromosomes diploides.

Selenoproteïnes
El seleni és un micronutrient poc abundant a la natura i essencial tant pels animals com per altres organismes. El paper del Seleni s'ha atribuït principalment a la seva presència en les Selenoproteïnes formant part de l'aminoàcid Selenocisteïna (Sec), l'aminoàcid número 21. En tant que el Seleni forma part de la composició de les Selenoproteines, es pot dir que és responsable de processos cel·lulars antioxidants, de metabolisme tiroideu i d'immunitat. El fet de que es trobi localitzat dins de les Selenoproteïnes i no lliure per la cèl·lula permet evitar la toxicitat del micronutrient. S'ha comprovat que tant el dèficit com l'excés d'aquest micronutrient comporta efectes perjudicials per a la salut. El dèficit de Seleni, per exemple, es pot traduir en la malaltia de Keshan, caracteritzada per una necrosis a nivell de miocardi que comporta una disfunció cafdíaca. A més a més, la suplementació de seleni s'ha correlacionat amb la prevenció d'alguns dels càncers humans.
El seleni és un micronutrient poc abundant a la natura i essencial tant pels animals com per altres organismes. El paper del Seleni s'ha atribuït principalment a la seva presència en les selenoproteïnes formant part de l'aminoàcid selenocisteïna (Sec), l'aminoàcid número 21. En tant que el Seleni forma part de la composició de les selenoproteïnes, es pot dir que és responsable de processos cel·lulars antioxidants, de metabolisme tiroideu i d'immunitat. El fet de que es trobi localitzat dins de les selenoproteïnes i no lliure per la cèl·lula permet evitar la toxicitat del micronutrient. S'ha comprovat que tant el dèficit com l'excés d'aquest micronutrient comporta efectes perjudicials per a la salut. El dèficit de Seleni, per exemple, es pot traduir en la malaltia de Keshan, caracteritzada per una necrosis a nivell de miocardi que comporta una disfunció cardíaca. A més, la suplementació de seleni s'ha correlacionat amb la prevenció d'alguns dels càncers humans.

Les selenoproteïnes són proteïnes que incorporen en la seva seqüència una o més selenocisteïnes, un aminoàcid descrit com anàleg de la cisteïna. La única diferència que presenta la selenocisteïna respecte la cisteïna recau sobre la presència d'un àtom de seleni en lloc de l'àtom de sofre, tots dos presenten propietats químiques similars ja que es localitzen en la mateixa columna de la taula periòdica. Per aquest motiu es poden trobar en algunes espècies homòlegs en cisteïna, selenoproteïnes on la selenocisteïna està substituïda per una cisteïna.
Síntesi de les Selenoproteïnes
El codó que codifica per la selenocisteïna és l'UGA, triplet que normalment correspon a un stop codon. Per tant, en una mateixa espècie tenim un mateix codó UGA amb dues interpretacions diferents. Per tal de que aquest nou significat del triplet sigui possible es necessita una maquinària extra i específica. El ribosoma, per tal de diferenciar un stop codon d'un codó codificant per a la selenocisteina, caldrà que es trobi amb un SECIs (SelenoCysteine Insertion Sequence), element tridimensional de l'estructura secundària de l'RNA. La forma del SECIs consisteix en un loop que en funció del regne pren una localització o un altra: en bacteris es troba en regions codificants dels gens, mentre que en archae i eucariotes es troba en l'extrem 3'UTR.
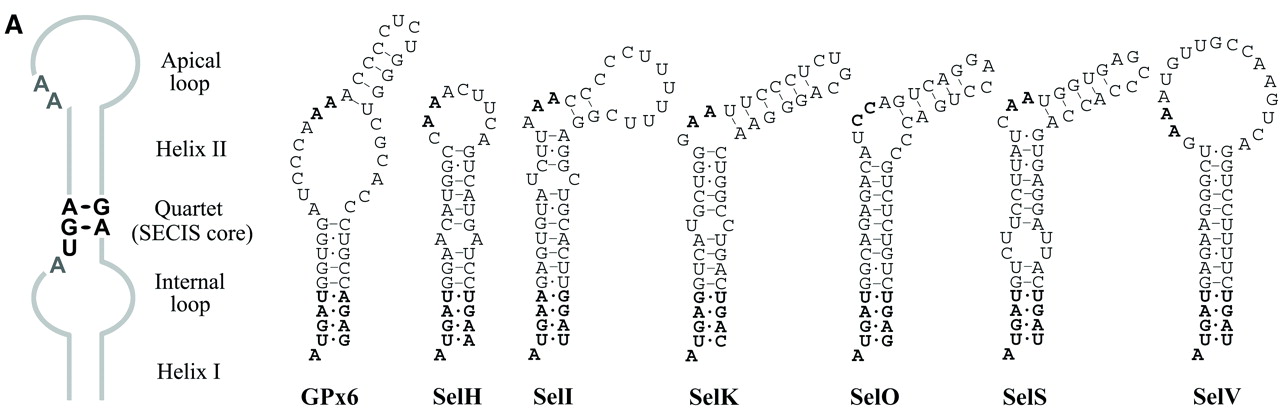
La funció del SECIs és unir SBP2 (SECIs Binding Protein), proteïna que recluta eEFSec, un factor d'elongació específic de selenoproteïnes que selecciona el tRNA que està unit a SECIs per tal d'apropar-lo al codó UGA del mRNA i així facilitar la incorporació de la selenocisteïna. Fins que el ribosoma no reconegui un codó de terminació no acabarà la traducció del mRNA. Hi ha altres proteïnes que participen específicament en la síntesi de la selenocisteïna, són la selenofosfat sintetasa (SPS1 i SPS2), la selenocisteïna sintasa (SLA/LP), la fosfoseril tRNA quinasa (PTSK) i la proteïna associada a selenocisteïna (SECP43).

Cal considerar que la identificació automatitzada de les selenoproteïnes es veu limitada per la consideració sistemàtica dels codons UGA com a stop codons i no com a triplets codificant per a una selenocisteïna. A primera vista, un bon mètode per discriminar entre aquestes dues "lectures" del mateix codó podria ser buscar els SECIS, si no fos perquè aquests no presenten un patró conservat de la seqüència. Tot i així, es coneix que hi ha SECIS que conserven el quartet AUGA i altres que contenen altres quartets, anomenats canònics i no canònics respectivament. La millor estratègia per identificar aquests elements SECIS recau sobre la cerca de seqüències amb la capacitat de plegament i de formació de l'estructura característica del SECIs: loop-helix-loop. Finalment, la estratègia òptima per predir la presència de selenoproteïnes en un genoma es basa tant en la informació sobre els exons del gen d'estudi com en la identificació dels elements SECIS.