El món del Seleni
1. El Seleni a la natura
El Seleni (Se) és un element essencial per totes les formes de vida, tant eucariotes com procariotes. Tot i no ser molt abundant, forma part de diverses vies metabòliques principals, incloent el metabolisme tiroideu, sistemes antioxidants i sistema immunitari. Una expressió subòptima d'enzims amb contingut de seleni pot donar lloc a diverses disfuncions, d'altra banda, un excés de seleni és tòxic (selenosi).
En el cas dels animals, dels eucariotes i dels protists, el seleni no està lliure sinó que es troba en les cèl·lules en forma de selenoproteïnes.[1]
2. Les Selenoproteïnes
Les selenoproteïnes són proteïnes que incorporen un aminoàcid anomenat selenocisteïna (aminoàcid vint-i-u). Aquest aminoàcid és present en la major part d'espècies (archaea, procariotes i eucariotes), però el contingut de selenoproteïnes és molt variable en els diferents organismes, fins i tot hi ha organismes que no tenen aquest tipus de proteïnes. [2]
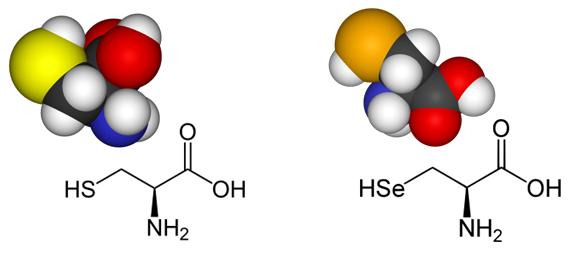
La selenocisteïna (Sec) és un anàleg de la cisteïna (Cys) que en comptes de tenir un àtom de sofre (S), conté un àtom de seleni (Se). Aquests dos elements tenen una reactivitat semblant i presenten unes propietats químiques similars.
Aquest aminoàcid ve codificat pel codó UGA, que normalment s'interpreta com stop codon. En eucariotes, aquesta recodificació és possible per l'estructura secundària que adquiereix l'mRNA gràcies a un motiu en cis situat al seu 3'-UTR, anomenat element SECIS
3. Biosíntesi de Selenoproteïnes
No hi ha cap codó que codifiqui per Sec, per tant, la selenocisteïna ha estat inserida en polipèptids per una recodificació del codó UGA, un codó STOP. És el primer cas en què un codó en una mateixa espècie té dos productes diferents.
3.1. Síntesi de l'aminoàcid Selenocisteïna
Durant la biosíntesi de de l'aminoàcid selenocisteïna, uns tRNAs únics que tenen codons complementaris UCA són aminoacilats amb serina formant el seril-tRNA. Aquest tRNA pot ser fosforilat per la fosfoseril tRNA quinasa (PSTK). Paral·lelament, l’enzim selenofosfat sintetasa (Sps2 en eucariotes) sintetitza el monoselenofosfat a partir de seleni ingerit per la dieta, que serà el donador de Se. Seguidament es transfereix del selenofosfat al seril-tRNA formant SeC-tRNASeC.
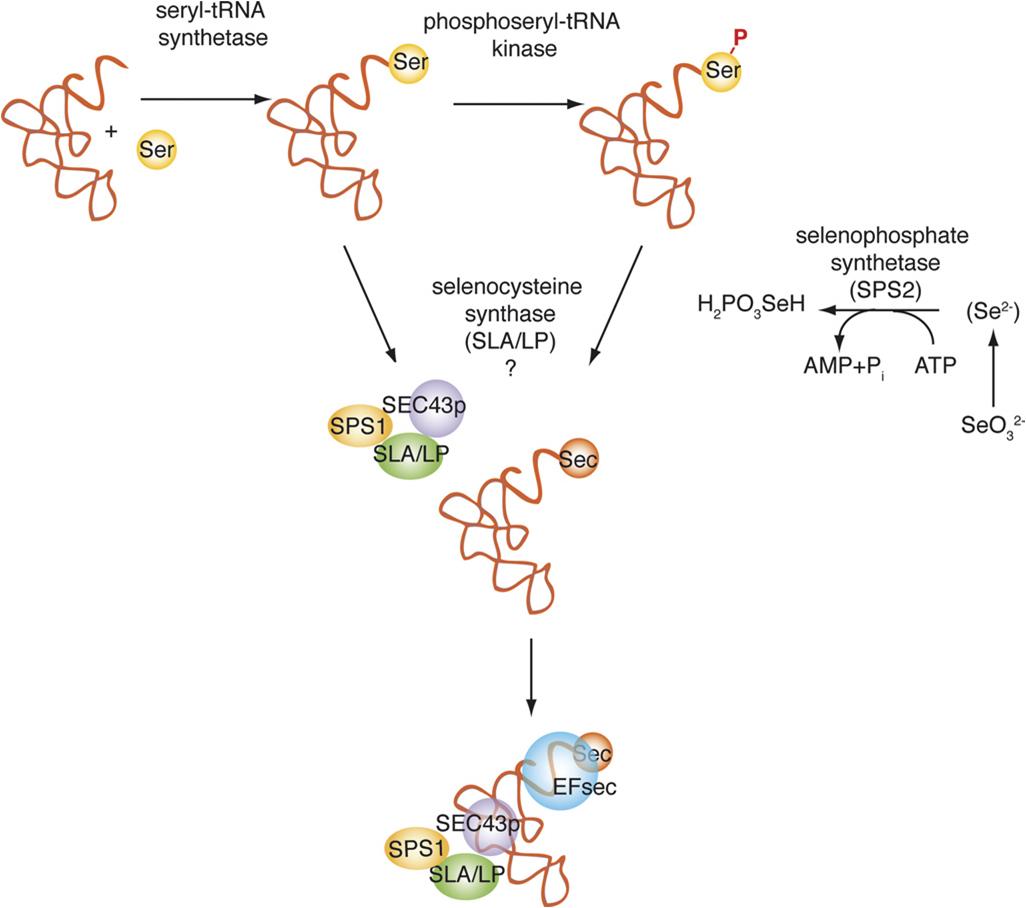
3.2. Inserció de la Selenocisteïna a l'estructura primària
Un cop tenim el tRNA de l'aminoàcid selenocisteïna, cal la seva incorporació a les cadenes peptídiques durant traducció. La recodificació del codó UGA, codó stop normalment, com a codó per la selenocisteïna és mediada per una estructura tridimensional en l’extrem 3’ no traduït (UTR) dels gens de les selenoproteïnes. Aquesta estructura rep el nom de SElenoCystein Insertion Sequence (SECIS Element).
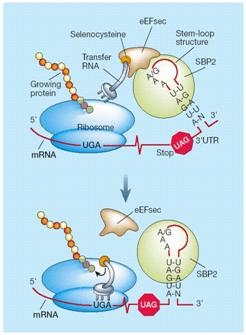
Aquest element recluta una proteïna anomenada SECIS Binding Protein (SBP2), la qual s’uneix a un factor d’elongació específic de selenoproteïens (Efsec). Efsec apropa el tRNA de la selenocisteïna (tRNASec) al codó UGA del mRNA de les selenoproteïnes i en determina la seva traducció a selenocisteïna. La traducció del mRNA continua fins que el ribosoma reconeix el següent codó de terminació. D’aquesta manera, el ribosoma, que normalment quan trobava l’UGA acabava la traducció, el que fa és reconèixer l’UGA per un codó amb l’aminoàcid selenocisteïna i continua la lectura fins el següent codó de terminació, que pot ser qualsevol dels tres codons (UAG, UAA o UGA).[3][4]
4.Recerca de Selenoproteïnes
La utilització excepcional del codó UGA per codificar selenocisteïnes (Sec), fa molt difícil la identificació de les selenoproteïnes ja que pràcticament tots els programes bioinformàtics assumeixen que aquest codó codifica per una senyal de terminació de la traducció. És per aquest motiu que la identificació de selenoproteïnes requereix la utilització d’altres mètodes. A més, sense supervisió humana normalment resulta molt difícil caracteritzar el selenoproteoma d’un determinat genoma.
Per aquesta raó és important la recerca de selenoproteïnes en diferents genomes seqüenciats, tant de protistes com d'eucariotes. Concretament el nostre projecte consisteix en la recerca en catorze genomes de protistes de les següents families de selenoproteïnes:
- Familia GPx
Familia de glutatió peroxidases. Aquestes proteïnes són la principal defensa enzimàtica antioxidant en humans: participen en l'eliminació del peròxid d'hidrogen, modulen la síntesi d'eicosanoides modificant la inflamació i disminueixen el dany oxidatiu de lípids, lipoproteïnes i DNA. S'han descrit vuit proteïnes homòlogues per aquesta familia, cinc de les quals són selenoproteïnes:
- GPx1: cel·lular o gàstrica - GPx2: gastrointestinal - GPx3: extracel·lular o plasmàtica - GPx4: associada a la membrana mitocondrial - GPx6: epiteli olfactori i teixit embrionari
Les GPx5, GPx7 i GPx8 no han estat descrites com a selenoproteïnes.
- Familia FrnE
FrnE, també coneguda com Dsba (disulfide bond formation protein A) és una selenoproteïna de la superfamilia de les tioredoxines. En un primer moment, només s'havia descrit en múltiples organismes procariotes i microorganismes de comunitats marines, entre d'altres. Recentment, aquesta selenoproteïna ha estat descrita en organismes multicel·lulars com Ciona intestinalis.
5.Homòlegs i paràlegs de Selenoproteïnes
A vegades, l’ortòleg d’una selenoproteïna conté una cisteïna enlloc d’una selenocisteïna. Molt sovint trobem que la mateixa proteïna en una espècie determinada és una selenoproteïna, en canvi en una altra espècie, en la mateixa posició, en comptes d’una selenocisteïna hi ha una cisteïna, per tant tenim un homòleg en cisteïna. Fins i tot, dins de la mateixa espècie trobem un gen amb selenocisteïna i una còpia del mateix amb una cisteïna (paràleg). Normalment les selenoproteïnes tenen només un aminoàcid selenocisteïna, però hi ha excepcions i algunes en tenen més d’un.
El Regne dels Protistes
1.Introducció al Regne dels Protistes
1.Introducció al Regne dels Protistes
Els protistes són un grup molt divers de microorganismes eucariotes. Tradicionalment, eren considerats un regne independent degut a l'àmplia varietat de característiques que dificultava incloure'l en cap altre. Actualment, però, es classifiquen filogenèticament.. Dins d'aquest regne històric, sel's classificava en tres grups diferents segons les semblances amb altres regnes:
- Protozous unicel·lulars semblants als animals.
- Protofitins semblants a les plantes.
- Mixomicets i oomicets semblants als fongs.
Tenen organització i estructures biològiques relativament senzilles respecte la resta d'eucariotes (fongs, animals, plantes) ja que poden presentar estructura unicel·lular, pluricel·lular o colonial sense arribar a formar teixits especialitzats.
Viuen gairebé en qualsevol medi que contingui aigua líquida. Amb un estil de vida molt similar al dels procariotes, la majoria són autòtrofs i aerobis. Molts protists com ara les algues, són fotosintètics i productors primaris essencials dels ecosistemes, mentre que altres com els cinetoplàstids i apicomplexos causen malalties als humans com ara la malària.
2.Els protistes que estudiem
