






| |
|
|
|
|
|

Introducció
La proteïna CREB (cyclic AMP-responsive element-binding
protein) és un factor de transcripció clau que estimula
l’expressió de nombrosos gens en resposta a
factors de creixement, hormones, neurotransmissors, fluxes
iònics i senyals d’estrès. Aquests elements extracel·lulars
activarien un conjunt de missatgers secundaris que transduirien la senyal via rutes quinasa
fins l’activació de CREB, que té localització nuclear. Aquesta
activació es porta a terme mitjançant la fosforilació de la serina 133
d’aquest factor. Tant CREB com els seus paràlegs ATF1 i CREM actuen formant
dímers i activant la transcripció de gens corresponents a un ampli ventall de
processos biològics, incloent-hi el metabolisme glucídic i la supervivència
cel·lular i complexes funcions neuronals com la memòria i l’aprenentatge.
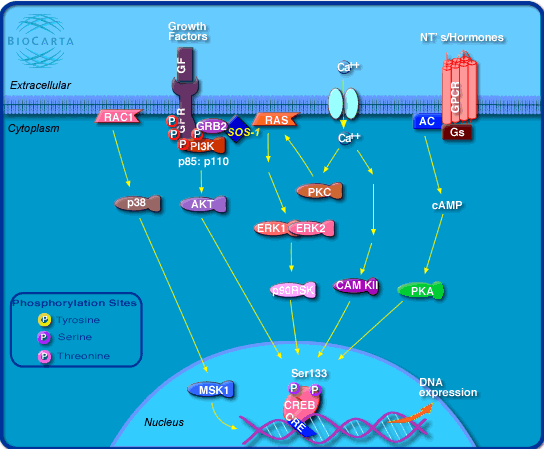 Figura 1: vies d'activació de CREB
Figura 1: vies d'activació de CREB
Els membres de la familia CREB s’uneixen al DNA per mitjà d’un domini
proteic bàsic anomenat leucine zipper (bZIP). Aquest motiu és
capaç de reconèixer una seqüència palindròmica curta present
en el genoma: l’element CRE (cAMP response element).
Aquesta seqüència es localitza freqüentment a la regió promotora proximal
al lloc d’inici de transcripció (TSS:
Altres estudis indiquen que CREB està unit constitutivament als motius CRE fins i tot en
absència d’agonistes. Aquesta unió es produeix molt sovint molt a prop de
les capces TATA del mateix promotor. Per aquest motiu, la unió de CREB al
promotor sembla estimular interaccions cooperatives amb altres factors de transcripció
per tal de facilitar el reclutament dels complexes RNA polimerasa II.
Tornar a Taula de continguts
Objectius
L’objectiu del nostre estudi és analitzar la possible cooperació
funcional entre els factors d’unió als motius CRE i TATA del DNA. Volem
estudiar aquesta interacció putativa comparant la distribució dels dos motius
en diferents seqüències corresponents a diferents regions gèniques humanes.
Tornar a Taula de continguts
Materials i Mètodes
Per realitzar el nostre estudi hem utilitzat dos fitxers que contenen seqüències
corresponents a diferents regions gèniques en format FASTA:
Per tal d’analitzar la distribució dels motius CRE i TATA sobre aquestes
seqüències, hem utilitzat dues expressions regulars que defineixen la
seqüència consens de cadascun dels motius. Degut a que els elements de
resposta són seqüències relativament curtes, hem precisat les expressions
per tal de limitar la búsqueda sobre les seqüències. Així, les
expressions regulars utilitzades en format Perl són les següents:
Hem realitzat la búsqueda d’aquests motius sobre les seqüències
mitjançant el programa motius.pl, descrit detalladament a la secció PROGRAMA. Els
resultats obtinguts al fitxer de sortida s’han processat i representat en forma
d’histogrames a partir del programa SPSS v11.0.
Tornar a Taula de continguts
Resultats
 |
 |
|
 |
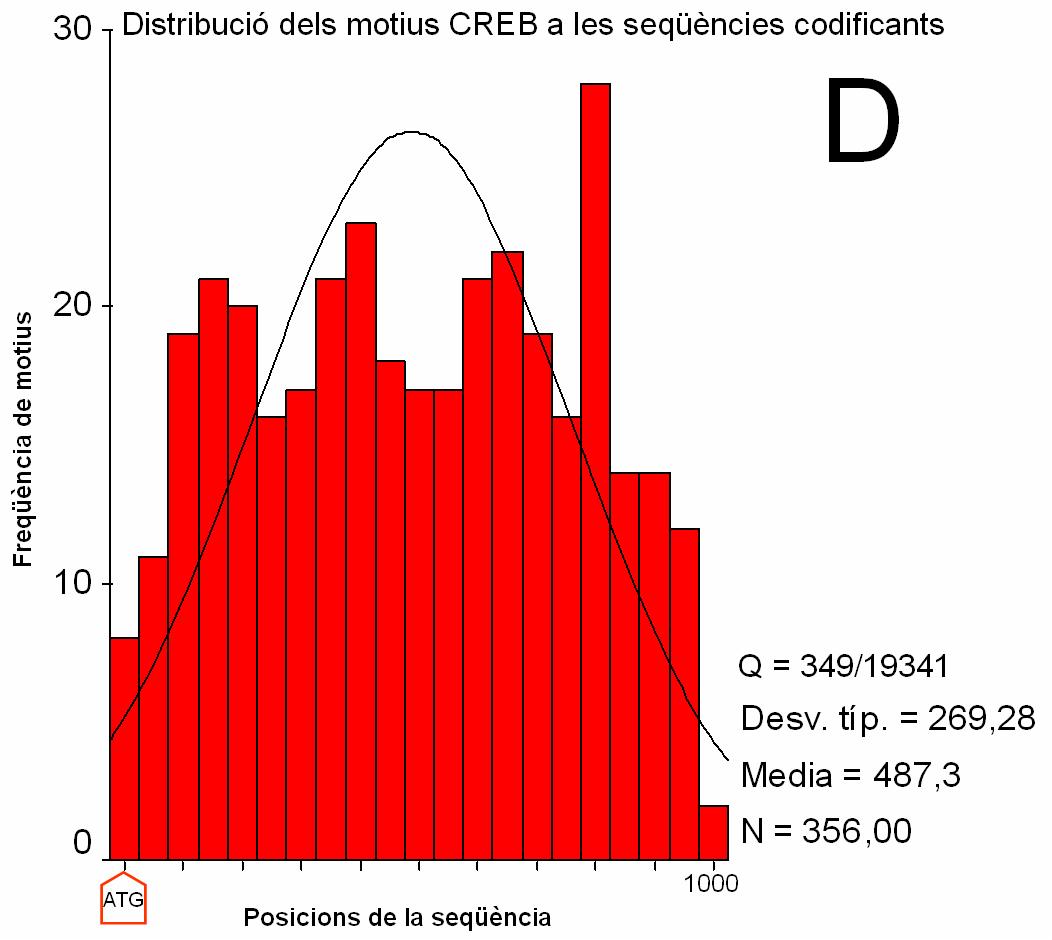 |
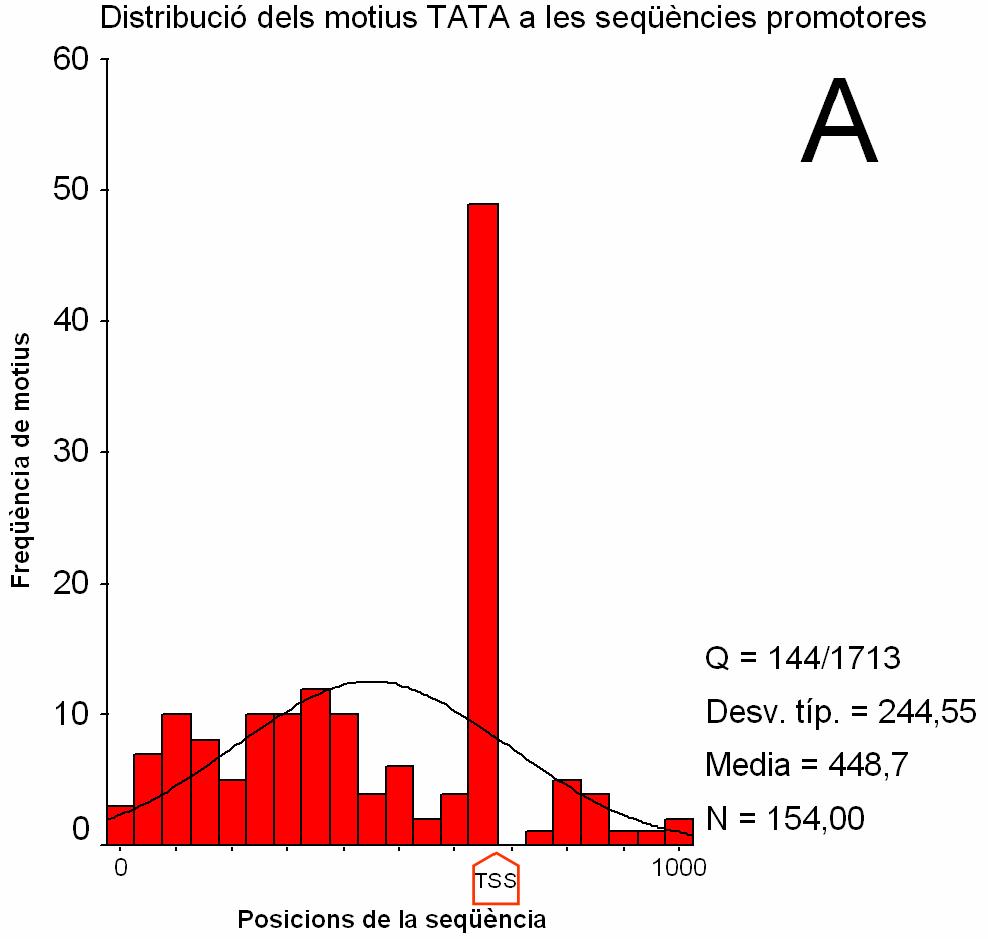
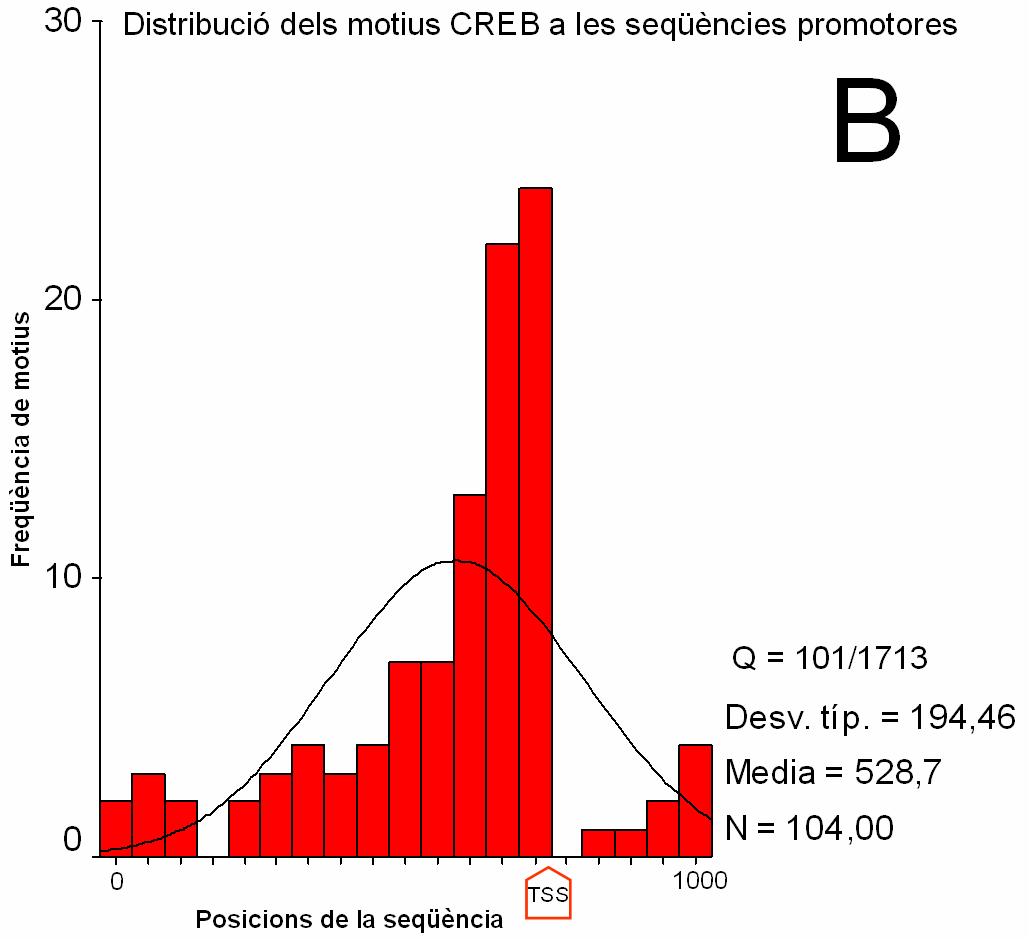
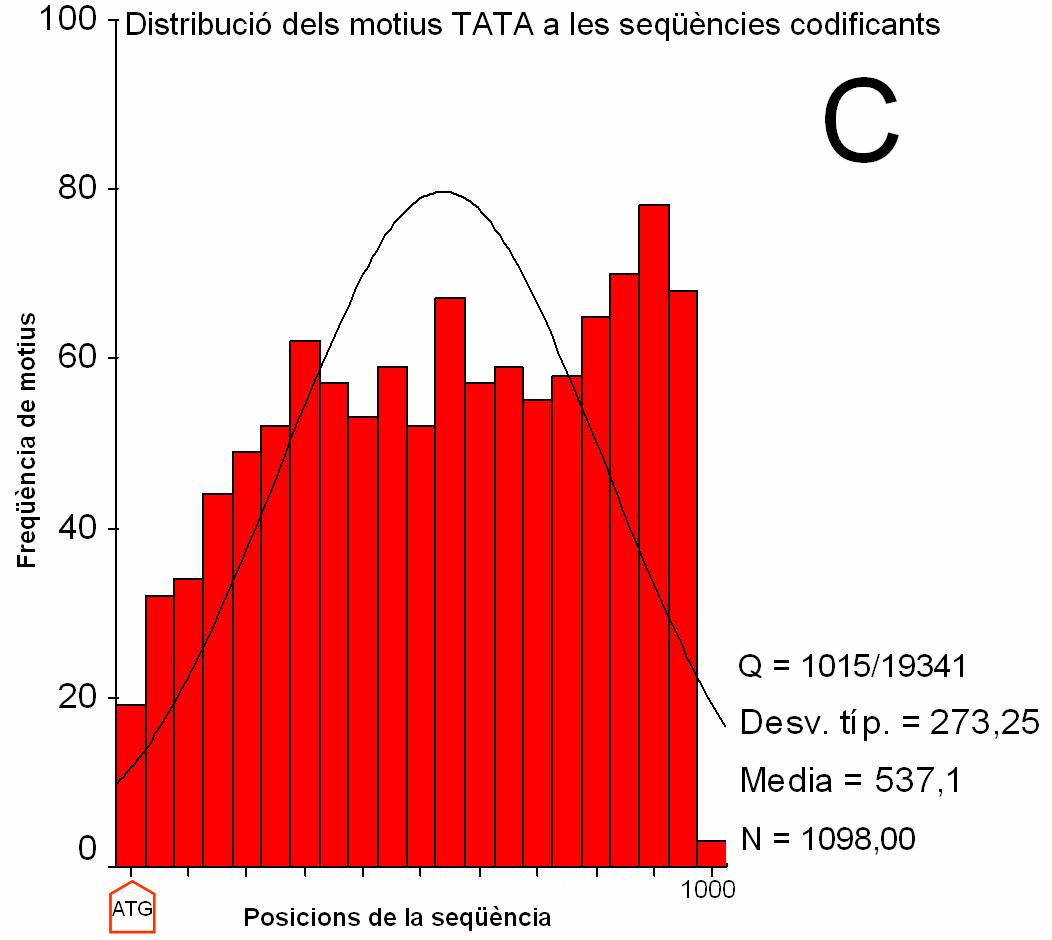
Figura 2.
Distribució dels motius CRE i TATA en seqüències
promotores i codificants (clica per veure els gràfics ampliats).
(A) i (B) Histograma que recull la freqüència de match registrada a cada intèrval de posició de les seqüències promotores. Els intèrvals són de 100 nucleòtids en A i 5 nucleòtids en B. S’indica amb una fletxa la posició del TSS. La posició 0 a l’eix d’abcises correspon al nucleòtid –674 respecte el TSS. La posició 1000 correspon al nucleòtid +325. (C) i (D) Histograma que recull la freqüència de match registrada a cada intèrval de posició de les seqüències codificants. Els intèrvals són de 100 nucleòtids. La posició 0 correspon a l’ATG (indicat amb una fletxa). Q: nombre de seqüències amb match respecte el total de seqüències. Desv. típ: desviació estàndard Media: mitjana N: nombre total de match |
