INTRODUCCIÓN
Hemos utilizado nuestro programa (rastreador.pl) para buscar las secuencias consenso de unión de TATA (TATAA) y CREB (TGACGTCA) en dos ficheros, uno correspondiente a secuencias promotoras y otro correspondiente a secuencias codificantes.
A continuación utilizamos el paquete estadistico R para analizar la frecuencia con que aparece cada motivo en las distintas posiciones de las secuencias. Presetamos los datos en los histogramas que se muestran a continuación (click para ampliar las imágenes)
|
|
TATA EN REGIONES CODIFICANTES
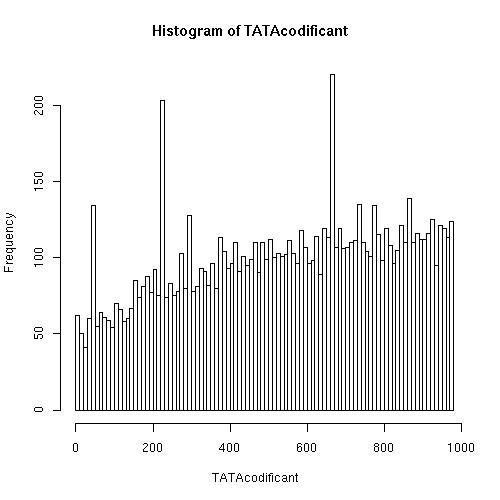
Hemos corrido en nuestro programa el fichero "hs_cod.fasta" correspondiente a varias secuencias codificantes a partir del codón de inicio de traduccion ATG.
El programa indica que le hemos introducido 19341 secuencias y ha encontrado el motivo de unión a TATA (TATAA) en 6633 de ellas.
Esto supone que el programa ha encontrado posibles dianas de union de TATA en el 34' 3 % de las secuencias introducidas. El procentaje es casi idéntico al de regiones promotoras (34' 7 %), con la gran diferencia de que en promotoras se concentra más entorno a una posición y en codificante aparecen a lo largo de toda la secuencia.
Teoricamente no deberíamos esperar encontrar una amplia distribución de este motivo en secuencias codificantes ya que su función esta restringida a la regulación de la transcripción situándose entre las posiciones -25 y -35 respecto al TSS en las regiones promotoras. Si pudiésemos realizar un estudio comparativo entre humano y otro mamífero como por ejemplo un ratón, veríamos conservación en las posiciones en que encontramos las dianas de TATA en los promotores pero no en DNA codificante, ya que estas no tienen una razón para conservarse evolutivamente (especialmente nos fijariamos en los intrones). A pesar de todo vemos una amplia distribución de este motivo, sin ninguna posición predominante sino que aparece a lo largo de todo el DNA codificante. Pensamos que estas dianas se dan simplemente por azar (nos parece fácil que se de al ser un motivo de solamente 5 bases).
|
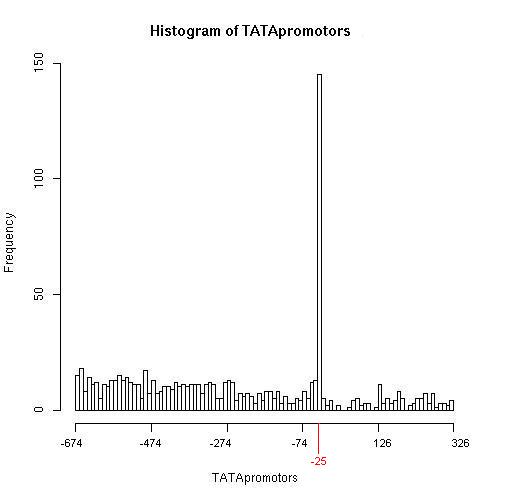 FIGURA2 FIGURA2 |
TATA EN REGIONES PROMOTORAS
Hemos corrido en nuestro programa el fichero "hs_EPD.fasta" correspondiente a varias secuencias de genes humanos desde la posicion -674 respecto al inicio de transcripción (TSS) hasta la 375 (secuencias que corresponden por tanto al promotor proximal).
El programa indica que le hemos introducido 1713 secuencias y ha encontrado el motivo de union a TATA (TATAA) en 595 de ellas.
Esto supone que el programa ha encontrado posibles dianas de unión de TATA en el 34' 7 % de las secuencias introducidas.
En aproximadamente 150 de las 595 secuencias en las cuales hemos encontrado la secuencia consenso TATAA ésta se situa 25 pares de bases por encima del inicio de la transcripción (-25) como se observa en el histograma.
Esta aproximación teórica se corresponde con la información que conocemos de la distribución de la caja TATA en las regiones promotoras de diferentes genes (concretamente ocupa un espacio que va desde la posición -35 a -25), reforzando la validez de nuestra aproximación a la distribución de diferentes motivos de union al DNA mediante técnicas informáticas basadas en el rastreo de secuencias consenso.
|
|
|
CREB EN REGIONES CODIFICANTES
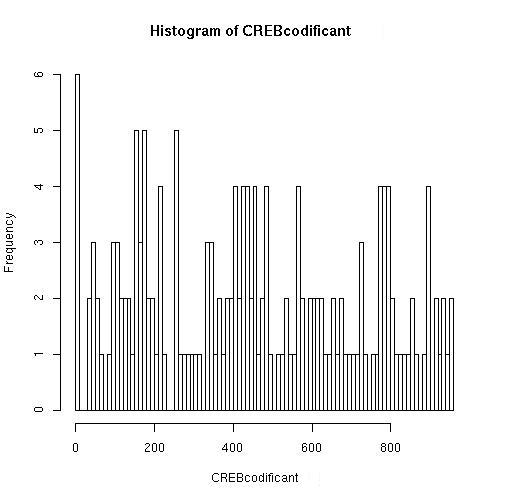
Hemos corrido en nuestro programa el fichero "hs_cod.fasta" correspondiente a varias secuencias codificantes a partir del codón de inicio de traducción ATG.
El programa indica que le hemos introducido 19341 secuencias y ha encontrado el motivo de unión a CREB (TGACGTGA) en 172 de ellas.
Esto supone que el programa ha encontrado posibles dianas de unión de CREB en el 0 ' 889 % de las secuencias introducidas. Comparando con el resultado de las dianas de CREB en regiones promotoras (4' 14%) vemos una notable disminución del porcentaje de secuencias en las que aparece el motivo de unión a CREB. Como en el caso de TATA, este motivo no tiene sentido en DNA codificante por lo que encaja con que lo hayamos observado en menos secuencias. Además en regiones codificantes nuevamente como en el caso de TATA la distribución de las dianas de CREB es uniforme a lo largo de toda la secuencia con variaciones poco significativas, así que nuevamente atribuimos al azar la presencia de estas dianas.
|
|
|
CREB EN REGIONES PROMOTORAS
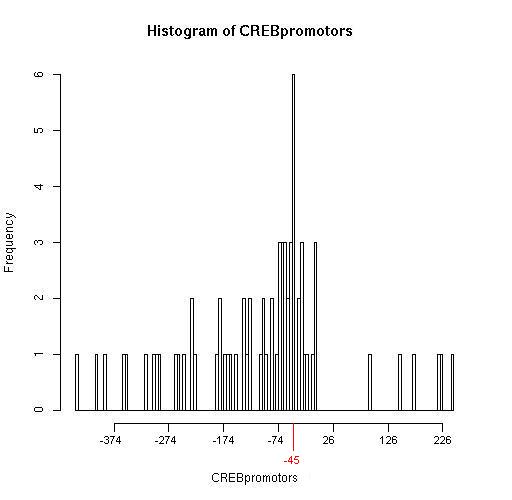
Hemos corrido en nuestro programa el fichero "hs_EPD.fasta" correspondiente a varias secuencias de genes humanos desde la posición -674 respecto al inicio de transcripción (TSS) hasta la 375 (secuencias que corresponden por tanto al promotor proximal).
El programa indica que le hemos introducido 1713 secuencias y ha encontrado el motivo de unión a CREB (TGACGTGA) en 71 de ellas.
Esto supone que el programa ha encontrado posibles dianas de unión de CREB en el 4' 14 % de las secuencias introducidas.
En aproximadamente 6 de las 71 secuencias en las cuales hemos encontrado la secuencia consenso TGACGTGA ésta se situa 45 pares de bases por encima del inicio de la transcripción (-45) como se observa en el histograma.
Asi, según nuestro programa las dianas del factor de transcripción CREB se situarían principalmente a distancias entre -100 y 0 respecto el TSS. Esto se corresponde con lo esperado según los artículos mencionados en la bibliografía.
En esta "figura" extraida del artículo "Genome..." se muestra la distribución de dianas de CREB conservadas en raton y humano y su localización se da principalmente entorno al 5'UTR.
También vemos una localización bastante similar de los motivos de unión de CREB y la caja TATA próximos al TSS. Esta proximidad no tiene porque suponer una localización conjunta en las regiones promotoras, puede tratarse simplemente de una posición preferente sobre la que actuarían CREB y otros factores de transcripción en la regulación de la expresión de diferentes genes, es posible encontrar en una secuencia solo el motivo de unión a CREB, solo el de TATA o que estén ambos o ninguno.
Volviendo a la información del artículo sabemos que existe una funcionalidad para la colocalización que se da en un 15% de casos, ya que la activación de CREB en respuesta a un aumento de los niveles intracelulares de AMPc va acompañada de un incremento en la expresión de aquellos genes cuyas secuencias promotoras tienen el motivo de unión para CREB y al mismo tiempo la caja TATA.
|
En el estudio anterior hemos concluido que la distribución de CREB y TATA en las regiones codificantes no respondía a ningún patrón significativo, es decir, era consecuencia del azar con que estos motivos se hayan presentes en las diferentes secuencias.Para reforzar esta explicación probamos el programa con expresiones regulares de longitud equivalente escogidas al azar. Los resultados los mostramos en el siguiente enlace.