ANŔLISI DE FAMÍLIES DE PROTEĎNES
PROTEĎNA LIFEGUARD
Introducció
Aquest treball caracteritza una família de proteďnes, partint d'una
seqüčncia problema (Lifeguard Q8K1F6 de Mus musculus). En primer
lloc, analitzarem les seqüčncies de proteďnes que presentin un bon
alineament amb la nostra proteďna (Programes: Swissprot, BLASTP, ClustalW,
Pfam). Seguidament, estudiarem les relacions filogenčtiques entre les
proteďnes escollides tot relacionant les funcions respectives (Programes:
ClustalW (Pasteur i EBI), TreeView).
I per últim, estudiarem els gens codificants pels membres d'aquesta
família proteica (cromosoma, exons) per humŕ i ratolí (Programes:
ENSEML, NCBI).
Entrem la nostra proteďna a la base de dades de Swissprot.
Hi trobem informació sobre funcions, seqüčncia d'aminoŕcids en format FASTA i
format TrEMBL, origen, referčncies...
Prenem la seqüčncia en format FASTA i fem córrer el programa BLASTP. Aquest programa compara la proteďna entrada amb totes les de la base
de dades en parells (dos a dos) a l'atzar, és a dir, la proteďna problema amb una proteďna a l'atzar, després respecte una altra
proteďna també a l'atzar, i així successivament. Atribueix a cada alineament una puntuació ("score") i un valor d'esperança
("E-value").
BLASTP té un match amb altres bases de dades de dominis proteics (Pfam, COG).
En les bases de dades de dominis, la proteďna que entrem és comparada amb el
conjunt de proteďnes semblants que troba dins la base
de dades. Les seqüčncies d'aminoŕcids conservades són més
significatives que si usem BLASTP ja que estan
contextualitzades únicament dins el grup de proteďnes
més similars.
Domini conservat
Les barres en vermell indiquen la predicció del domini conservat per a la proteďna "Lifeguard Q8K1F6"
de Mus musculus, segons les bases de dades Pfam i COG. El domini UPF0005 pertany a una família proteica no caracteritzada
("Uncharacterized Protein Family") segons la base de dades Pfam. I segons la base de dades COG el
domini COG0670 és un domini de proteďna integral de membrana que interacciona amb FtsH.
Les dues bases de dades suporten que el domini de conservació es troba en l'extrem C-terminal de la
seqüčncia proteica, aproximadament entre els aminoŕcids 140 i 310.

Escollim les seqüčncies proteiques de diferents espčcies, inclosa la
proteďna Q8K1F6 amb funcions diverses que compleixin un "score" més alt (millor
alineament) i una esperança ("E-value") mŕxima de 0.001 (si superem aquest rang
els valors no són estadísticament significatius). Obtenim així una mostra
representativa de diferents espčcies per tal de relacionar-les
evolutivament.
| Proteďna |
espčcie |
score |
E-value |
| Lifeguard; Q8K1F6 |
Mus musculus |
545 |
e-154 |
| Lifeguard; NMP35 |
Rattus norvegicus |
521 |
e-147 |
Lifeguard; KIAA0950 protein |
Homo sapiens |
501 |
e-140 |
| NMDA receptor glutamate-binding chain; LAG protein |
Mus musculus |
177 |
3e-43 |
| NMDA receptor glutamate-binding subunit |
Rattus norvegicus |
176 |
6e-43 |
| NMDA selective glutamate receptor; CG814-PA |
Drosophila melanogaster |
164 |
3e-39 |
| NMDA receptor associated protein |
Caenorhabditis elegans |
149 |
e-34 |
| NMDA1; CG3798-PA |
Drosophila melanogaster |
148 |
2e-34 |
| agCP6292 |
Anopheles gambiae |
148 |
2e-34 |
| transmembrane protein OTMP |
Ovis aries |
78 |
4e-13 |
| homolog of Drosophila NMDA1 protein, putative |
Plasmodium falciparum |
77 |
7e-13 |
| NMDA receptor-like protein; CMLV006 |
Camelpox virus |
75 |
2e-12 |
| z-protein |
Homo sapiens |
72 |
2e-11 |
Mitjançant el programa ClustalW (EBI) realitzem l'alineament múltiple de les
seqüčncies proteiques escollides per tal de limitar l'alineament a la zona
conservada.
Aquest és el resultat.
Paral.lelament, hem cercat la seqüčncia del domini UPF0005 (Mus musculus) a la base de dades Prosite i
comprovem que realment coincideix amb la seqüčncia conservada deduďda a partir de l'alineament múltiple anterior (ClustalW).
FPAGHHEHFTTFSWDDQKVRRLFIRKVYTILLVQLLVTLAVVALFTFCDVVKDYVQANPGWYWASYAVFFATYLTLACCS
pfam:UPF0005 <================================================================
GPRRHFPWNLILLTIFTLSMAYLTGMLSSYYNTTSVLLCLVITALVCLSVTIFSFQTKFDFTSCQGVLFVLLMTLFFSGL
pfam:UPF0005 ================================================================================
LLAVLLPFQYVPWLHAVYAVLGAGVFTLFLAFDTQLLMGNRRHSLSPEEYIFGALNIYLDIIYIFTFFLQLYGTNRE
pfam:UPF0005 ===========================================================================>
Prenem les seqüčncies seleccionades prčviament per tal de relacionar-les
evolutivament en un arbre filogenčtic (pel mčtode de "Neighbour joining").
Els dos programes clau són: ClustalW (Pasteur) i TreeView. L'opció "bootstraping"
per l'arbre "NJ" calcula 1000 vegades l'alineament previ a l'atzar i dóna una mesura
de robustesa (nombre de vegades que es genera el mateix arbre), en el nostre cas,
dóna exactament el mateix arbre i amb una elevada robustesa (prňxima a 1000).
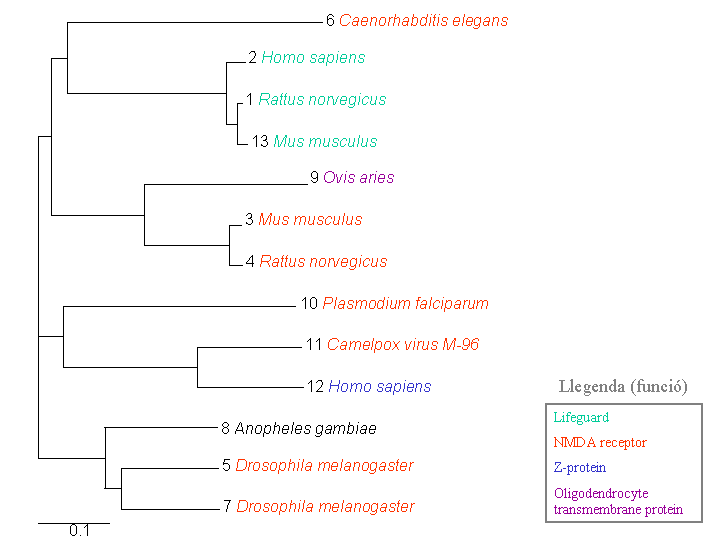
En l'arbre, observem una primera ramificació on s'agrupen d'una banda les lifeguards de
mamífer relacionades amb les NMDAs també de mamífer i de C. elegans; d'altra
banda una segona ramificació composta per Z-protein humana directament relacionada amb
Camelpox virus i secundŕriament amb P. falciparum,
espčcies que infecten i parasiten l'home respectivament; i un tercer cluster protagonitzat
per proteďnes d'insectes (NMDAs de Drosophila sp. i agCP6292 d'Anopheles
gambiae).
Comentarem les funcions de les diferents proteďnes. Com veiem a l'arbre,
les proteďnes a distŕncia filogenčtica més propera són les lifeguards (de
Rattus norvegicus, Homo sapiens i Mus musculus).
La proteďna lifeguard pot unir-se al receptor de Fas perň no regula l'expressió
de Fas ni interfereix amb la unió d'un agonista. Lifeguard és una molčcula
anti-apoptňtica (protegeix les cčl.lules de l'apoptosi induďda per Fas) (1).
La maquinŕria intracel.lular responsable de l'apoptosis és similar en totes les
cčl.lules animals. Aquesta maquinŕria depčn d'una família de proteases que
hidrolitza les proteďnes diana en el seu costat carboxil dels ŕcids aspŕrtics
(d'aquí el seu nom, c-caspasa). Les caspases són sintetitzades com a precursors
inactius (procaspases) que poden activar-se per l'esmentada ruptura. Així,
un cop desencadenada l'activació, té lloc una cascada de caspases (la qual
provoca la degradació de proteďnes estructurals, de factors de transcripció,
etc, acabant amb la mort cel.lular). L'activació de les procaspases pot ser
induďda des de l'exterior de la cčl.lula mitjançant l'activació de receptors
mortals presents a la superfície cel.lular. El lligand de Fas unit al
receptor Fas activa la cascada (2). Lifeguard és present a
les sinapsis de l'adult del Sistema Nerviós Central.
En humŕ, lifeguard (KIAA0950) s'expressa
en tots els teixits excepte melsa i placenta (1).
Els receptors NMDA
(N-metil-D-aspartat) són els típics receptors ionotrňpics (que formen un
canal iňnic) activats per l'aminoŕcid glutamat. El glutamat és el
neurotransmissor excitador més important en el SNC. Quan el receptor NMDA és activat,
augmenta la permeabilitat de la membrana plasmŕtica pel Calci. L'entrada de Calci
a l'interior cel.lular comporta innumerables efectes postsinŕptics. En repňs aquest
receptor estŕ bloquejat pel Magnesi (normalment present al medi extracel.lular).
La suma d'ambdues propietats (permeabilitat al Calci i bloqueig pel Magnesi) el
fan responsable de fenňmens de naturalesa plŕstica implicats en els processos
d'aprenentatge i memňria (3). El fet que Camelpox virus tingui
aquest receptor suggereix que l'ha adquirit per infecció d'una cčl.lula hoste,
possiblement d'Homo sapiens, per la seva proximitat evolutiva (reflectida
a l'arbre filogenčtic).
Z-protein és una proteďna eucariota integral de membrana relacionada amb GBP
(Glutamate Binding Protein), és a dir amb NMDA. Té 7 dominis transmembrana,
que pertanyen a la família BI1 (supressor apoptňtic de mamífers). La base de
dades GO prediu la seva funció: regulador de l'apoptosi.
La proteďna transmembrana d'oligodendrňcit (OTMP) també és una GBP. És una proteďna
integral de quatre hčlix amb un loop extern i amb les regions N i C-terminals
localitzades dins la cčl.lula. Es suggereix que OTMP és un receptor de
senyalització amb un paper en la mielinogčnesi (4).
La proteďna d'Anopheles gambiae és de funció desconeguda.
Tant les proteďnes lifeguard com els receptors NMDA contenen el domini BI1
(Bax inhibitor-1). El domini BI1 és similar a la família de proteďnes
intracel.lulars Bcl-2 (que contribueix a regular l'activació de les
procaspases). Alguns membres d'aquesta família (com el mateix Bcl-2 o
Bcl-XL) inhibeixen l'apoptosi bloquejant l'alliberament del citocrom C
(transportador d'electrons) del mitocondri (2).
Existeix una homologia significativa en la meitat C-terminal entre les proteďnes:
- Neural Membrane Protein 35 (NMP) de Rattus sp.
- Glutamate Binding Protein de Rattus sp.
- Drosophila NMDA receptor-associated protein
- dues proteďnes de funció desconeguda de C. elegans
demostrada per un article [PubMed]
[Free Full Text]
(1) trobat a la base de dades PubMed. Alhora, el contingut d'aquest article reflecteix
una correlació entre NMDA i l'expressió de Fas i el seu lligand (FasL), i
un important paper protector anti-apoptňtic de LFG implicat en aquesta correlació. S'ha
observat expressió de Fas i FasL després d'injectar NMDA a l'hipocamp de ratolí (5).
Malalties com leucčmia i Alzheimer (amb un component apoptňtic i el cromosoma 12q13 humŕ
implicat) l'expressió i modulació de LFG pot tenir un important ús terapčutic quan
la protecció davant de Fas és requerida.
Hem ampliat la informació d'aquest arbre amb altres proteďnes d'interčs (que
apareixen en el tercer bloc) i, a més a més, amb una seqüčncia d'un herpes
virus humŕ (relacionat amb Lifeguard Q8K1F6 en l'article: (6)).
En aquest article hi ha evidčncies d'homologia entre Lifeguard humana i la proteďna HCMVA
(pertanyent a la família US12) de l'herpesvirus 5 humŕ, suggerint un paper anti-apoptňtic.
Aquesta proteďna és integral de membrana, interacciona amb FtsH, i conté el domini
UPF0005 amb subdomini BI1.
En l'alineament múltiple
de l'article citat es reflecteix el domini conservat UPF0005 entre membres de la família
US12 d'Herpesvirus, lifeguard humana, i dues proteďnes humanes addicionals.
Mitjançant el programa ClustalW (EBI) realitzem un alineament múltiple de totes aquestes
seqüčncies i una posterior representació de les relacions evolutives entre les diferents
espčcies implicades:
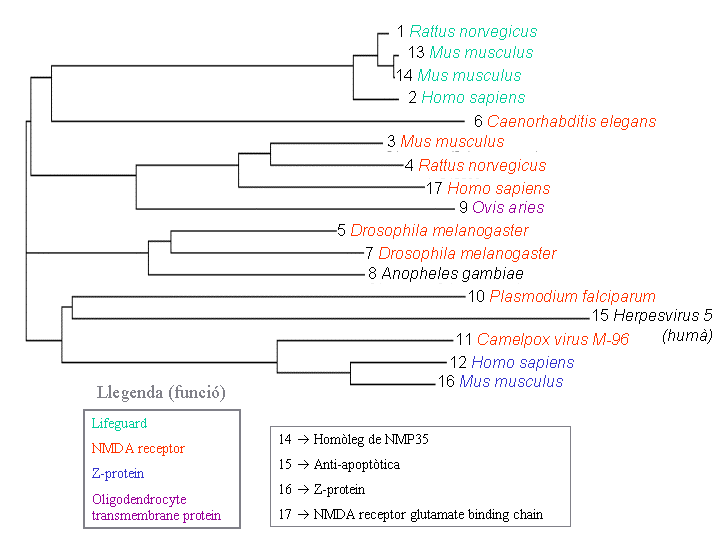
En aquest segon arbre, es manté la mateixa agrupació d'espčcies i funcions plasmada en el
primer arbre. Les quatre proteďnes (l'homňleg de NMP35 (14) i la Z-protein (16) de Mus musculus,
NMDA receptor glutamate-binding chain (17) de Homo sapiens, i una proteďna relacionada
d'Herpesvirus (15) humŕ) afegides s'integren en l'arbre sense desestructurar aquesta distribució.
D'aquesta manera, la proteďna número 14 es relaciona directament amb la lifeguard (13)
de la mateixa espčcie (Mus musculus); la número 16 directament amb la seva homňloga
Z-protein de Homo sapiens; la número 17 amb NMDA de rosegadors; i finalment, la
número 15 dins el mateix grup que P. falciparum i Camelpox virus
reflectint la possible adquisició de gens d'hostes humans, entre altres espčcies, al
llarg de l'evolució.
Entrant a l'ENSEMBL la seqüčncia FASTA de la nostra proteďna de partida
(Q8K1F6) per tal de córrer un BLASTP (córrer la proteďna contra la base de dades
de l'ENSEMBL). El resultat és que el millor alineament no és suficientment
significatiu per constatar que ENSEMBL conté la proteďna. Per tant, no podem
mostrar l'estructura exňnica de Q8K1F6.
Ja que no tenim un representant de
Lifeguard en Mus musculus, partim de la Lifeguard d'Homo
sapiens (KIAA0950). Prenem la seqüčncia humana en format FASTA i correm un
BLASTP contra les proteďnes de Mus musculus d'ENSEMBL. Així trobem la
seva proteďna homňloga en Mus musculus: Lifeguard Q9D6K4.
De la
mateixa manera, a partir de la seqüčncia FASTA de la proteďna NMDA receptor
glutamate-binding chain (Lag protein) de Mus musculus, trobem la seva
homňloga humana: NMDA receptor glutamate-binding chain (GRINA).
Segons la base de dades Pfam, totes les proteďnes de la taula anterior
comparteixen el domini conservat UPF0005. Els membres de la família UPF0005 es preveu
que contenen 7 dominis transmembrana i que comparteixen la regió compresa entre el
principi del 3er domini fins meitat del 4rt (1). La cerca per ENSEMBL de les
proteďnes de Mus musculus mostra el domini conservat BI-1 (Bax inhibitor-1),
mentre que per les proteďes de Homo sapiens mostra el domini conservat
UPF0005. La base de dades INTERPRO caracteritza BI-1 com a subdomini del domini
conservat UPF0005.
Segons la bibliografia (7), BI-1 és una proteďna integral de membrana
evolutivament conservada en mamífers, implicada en la supressió de l'apoptosi.
Conté múltiples segments transmembrana, com podem veure en totes les proteďnes de
la taula.
Trobem evidčncia d'Splicing Alternatiu en Z-protein i en Lifeguard
d'Homo sapiens. El primer cas, quatre transcrits generen quatre isoformes
diferents de la Z-protein. En el segon cas, es tradueixen dues isoformes de la
Lifeguard a partir de dos transcrits.
4.Referčncies
(1) Somia NV, Schmitt MJ, Vetter DE, Antwerp DV, Heinemann SF, Verma IM.
LFG: An anti-apoptotic gene that provides protection from Fas-mediated cell death.
Proc. Natl. Acad. Sci. USA. 1999 October 26;96(22):12667–12672.
(2) Alberts B, Johnson A, Lewis J, Raff M, Roberts K, Walter P.
Molecular Biology of the Cell. New York: Garland Science, 2002.
(3) Lerma J, Mora J, Prieto JS. Sinapsis monoaminérgicas y
peptidérgicas. In: Delgado JM, Ferrús A, Mora F, Rubia FJ. Manual de Neurociencia.
Madrid: Editorial Síntesis, 1998;233-234.
(4) Szuchet S, Plachetzki DC, Eaton KS.
Oligodendrocyte transmembrane protein: a novel member of the glutamate-binding protein subfamily.
Biochem Biophys Res Commun 2001 May 18;283(4):900-7.
(5) Shin SW, Park JW, Suh MH, Suh SI, Choe BK.
Persistent expression of Fas/FasL mRNA in the mouse hippocampus after a single
NMDA injection. Journal of Neurochemistry 1998;71,1773-1776.
(6) Holzerlandt R, Orengo C, Kellam P, Albŕ MM.
Identification of New Herpesvirus Gene Homologs in the Human
Genome. Genome Res 2002 Nov;12(11):1739-48.
(7) Xu Q, Reed JC.
Bax inhibitor-1, a mammalian apoptosis suppressor identified by
functional screening in yeast. Mol Cell
1998 Feb;1(3):337-46.
5.Autores
Marta Beltran Domingo (marta.beltran01@campus.upf.es)
Ariadna Echenique Vergés (ariadna.echenique01@campus.upf.es)