Bioinformàtica - 2on trimestre curs 2006/2007 - UPF
 L'any 2003 es complia el 50 anniversari del descobriment per part de
James Watson i Francis Crick de
l'estructura del DNA.
L'any 2003 es complia el 50 anniversari del descobriment per part de
James Watson i Francis Crick de
l'estructura del DNA.
Aquest descobriment de l'estructura del DNA confirmava la intuició de d'Erwin Schröedinger que l'any 1944 en el seu famňs llibre "What is life" aventurava que la matčria cromosňmica havia de ser "un cristall aperiňdic, constituďt per la successió d'un nombre petit d'elements isomčrics", la seqüčncia concreta dels quals és la responsable de la seva funcionalitat.
Els primers mesos de l'any 1953 foren força agitats; diversos grups competien pel descobriment de l'estructura del DNA. El 31 de desembre del 1952, Linus Pauling i Robert Corey van enviar un manuscript al Proceedings of the National Academy of Sciences on exposaven el seu model de l'estructura del DNA. La carrera semblava guanyada. Pauling, perň, escrivia "The structure that we propose is a three-chain structure, each chain being an helix". Watson i Crick van adonar-se de seguida que el model era incorrecte--entre d'altres raons perque era molt semblant a un model que ells havien considerat i abandonat previament--. Rosalin Franklin, que trevalla amb Maurice Milkins va arrivar tambe a la mateixa conclusi\'o. Van ser les difraccions de raigs X de Rosalin que van permetre a Watson i Crick deudir l'estructura correcta del DNA, a principis de Març d'aquell any, poc desprčs de la publicació de l'article de Pauling i Corey. L'article de Watson i Crick va ser publicat a Nature l'abril del 1953.
Podeu llegir més sobre aquests esdevenimets a la histňria de la Biologia Molecular escrita per Horace Freeland Judson, "The eigth day of creation".
 El mateix any, perň, es produďa tambč un altre fet important:
Frederick Sanger
obtenia la seqüčncia d'aminoŕcids de la insulina bovina-la primera
-proteďna de la qual es coneixia la seqüčncia. Sanger obtindria el
premi Nobel per aquest descobriment.
El mateix any, perň, es produďa tambč un altre fet important:
Frederick Sanger
obtenia la seqüčncia d'aminoŕcids de la insulina bovina-la primera
-proteďna de la qual es coneixia la seqüčncia. Sanger obtindria el
premi Nobel per aquest descobriment.
MALWTRLRPLLALLALWPPPPARAFVNQHLCGSHL VEALYLVCGERGFFYTPKARREVEGPQVGALELAG GPGAGGLEGPPQKRGIVEQCCASVCSLYQLENYCN
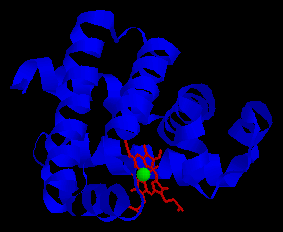
| 1959 determinació de la primera estructura tridimensional d'una proteďna. Es tracta de la mioglobina bovina, determinada mitjançant difracció de raigs X per Max Perutz i John Kendrew. | |

|
|
| 1957 Invenció dels llenguatges de programació d'alt nivell. Com ara el FORTRAN. Amb aquests llenguatges es possible escriure les instruccions per tal que un ordinador resolgui un determinat problema, sense cončixer com l'ordinador resol realment el problema. | |

|

|
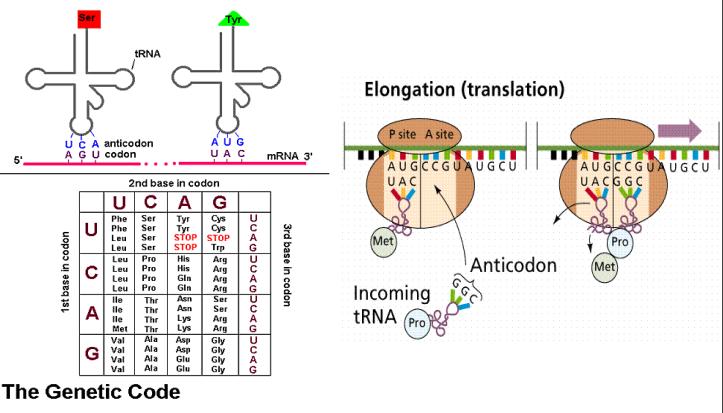
| principis dels 60. Desxiframent del codi genetic. Experiments de Korana, Brenner, Ochoa i altres permeten desxifrar el codi mitjançant el qual la seqüčncia de nucleňtids del DNA especifica la seqüčncia d'aminoŕcids de les proteďnes. | |
|
|
| anys 60s. els transistors substituiexen els tubs de buit en els ordinadors. En conseqüčncia, els ordinadors esdevenen més petits, rŕpids i econňmics. Cap a mitjans dels anys 60, la majoria de grans empreses processen la informació financera amb ordinadors digitals | |
|
|
|
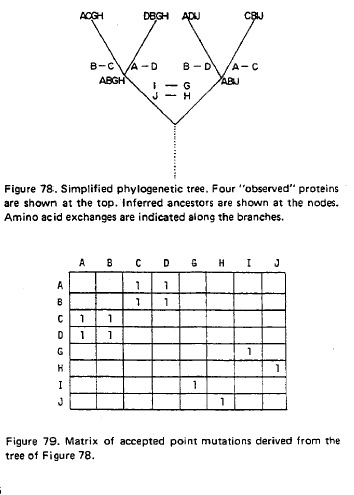
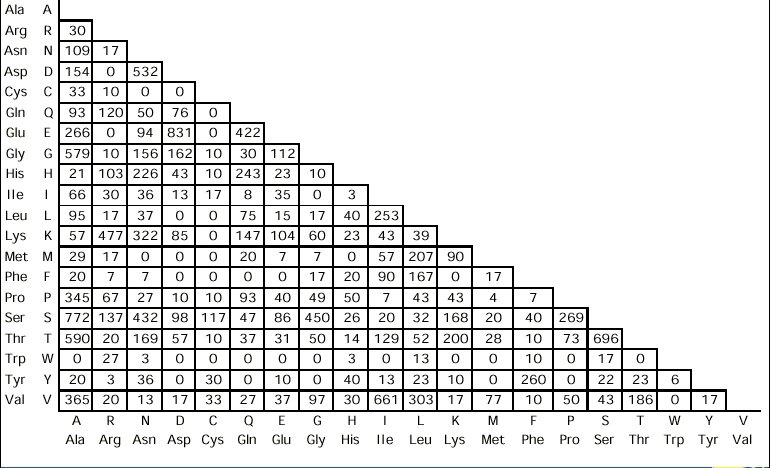
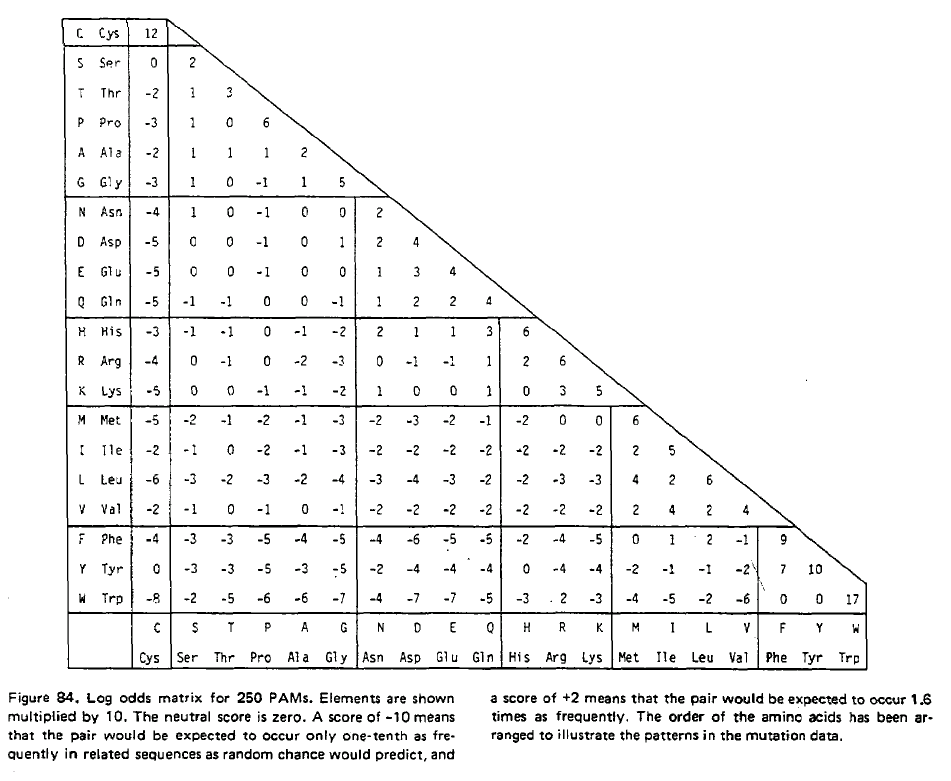
taula obtinguda de Douglas Brutlag Computational Molecular Biology Sequence Alignment que reprodueix els valors en l'article original de Dayhoff (1978).
Aquest taula contč en total 1572 canvis. Donat que aquests canvis ocorren a proteďnes relaciones estretament, corresponen a substitucions d'amionŕcids que no canvien essencialment la funció de la proteďna. Es per aixň que s'anomenen "Accepted Mutations", definides per Dayhoff com aquelles mutacions que són "acceptades" per la selecció natural. En aquesta taula, per exemple, es veu que hi ha 260 canvis entre l'aminoŕcid fenilalanina (Phe) i l'aminoŕcid tirosina (Tyr). Dayhoff, perň, estava interessada en calcular la probabilitat que un aminoŕcid mutés en un altre dins un determinat periode evolutiu. A partir d'aquesta taula i de les anomenades mutabilitats relatives de cada aminoŕcid (un valor proporcional a la rao del nombre de vegades que observe canvis en un aminoŕcid respecte el nombre total d'ocurrčncies d'aquell aminoŕcid),


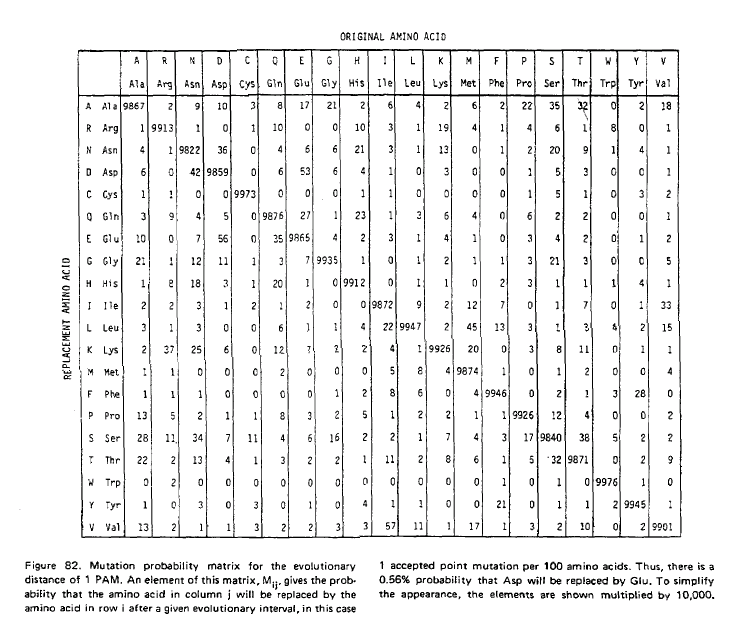
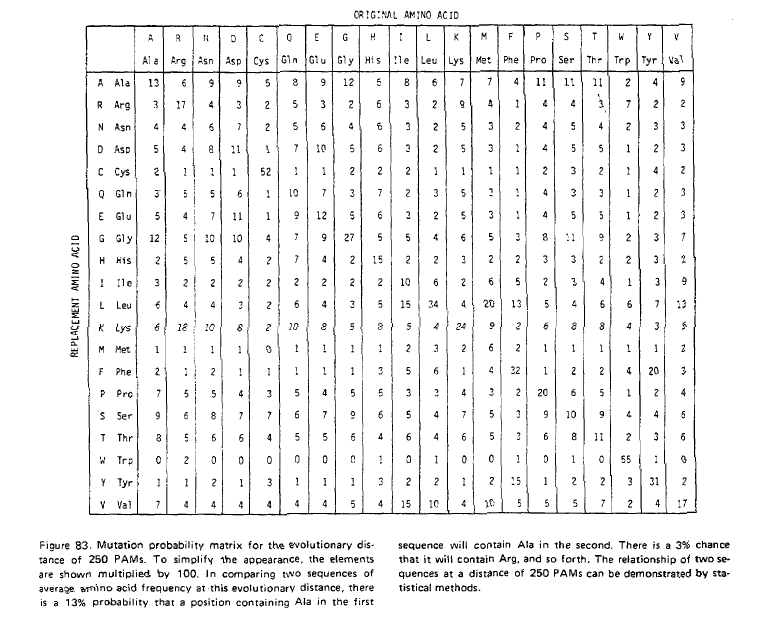
Normalment, hom calcula la matriu dels anomenats logaritmes de la rao de versemblança. En aquesta matriu, hom divideix les probabilitats observades de substitució d'una aminoŕcid per un altre per la probabilitat d'aquest aminoŕcid (es a dir, la probabilitat d'aparellar el primer amino acid pel segon a l'atzar) i, desprčs, calcula el logaritme d'aquesta raó. A la matriu resultant, hom calcula la mitjana dels valors corresponent als canvis recíprocs.