{kind=link}
Identificar open reading frames a partir de seqüències de DNA en les sis pautes de lectura possibles i traduir-les a proteïna. En el nostre cas hem analitzat part del genoma de l'herpesvirus associat a Sarcoma de Kaposi.
Per cada possible proteïna fem una cerca en BLAST per veure si és una proteïna ja coneguda, la seva funció i els homòlegs en altres organismes.
El nom d'Herpes prové del grec "Herpein" que significa amagar-se ja que aquest virus provoca una infecció crònica, latent i recurrent. Actualment s'han isolat aproximadament 100 herpesvirus. En totes les espècies animals en que s'han buscat se n'ha trobat almenys un. Es coneixen 8 herpes virus capaços d'infectar humans.
La família d'herpes virus es divideix en tres subfamílies:
INTRODUCCIÓ
| Alphaherpesvirinae: | |
|---|---|
| Simplexvirus | human herpesvirus 1, 2 (HSV-1, HSV-2) |
| Varicellovirus | human herpesvirus 3 (VZV) |
| Betaherpesvirinae: | |
| Cytomegalovirus | human herpesvirus 5 (CMV) |
| Muromegalovirus | mouse cytomegalovirus 1 |
| Roseolovirus | human herpesvirus 6, 7 |
| Gammaherpesvirinae: | |
| Lymphocryptovirus | human herpesvirus 4 (EBV) |
| Rhadinovirus | human herpesvirus 8 (HHV-8) |
Els herpes virus es caracteritzen per tenir genomes grans (>235kpb) que codifiquen per aproximadament 35 proteïnes. Aquestes proteïnes codifiquen per una gran varietet d'enzims relacionats amb el metabolisme dels àcids nucleics, la síntesi de DNA i procesament de proteïnes.
Característiques generals:
| Tamany | 180-200nm |
| Envolcall | Present |
| Tegument | Present |
| Càpside | Icosahèdrica, 162 capsomers hexagonals |
| Nucleocàpside | DNA envoltat per proteïnes |
| Genoma | Lineal,dsDNA |
| Replicació | Nuclear |
| Ensemblatge | Nuclear |
Tots els genomes dels herpes virus tenen una única regió llarga (UL) i una de curta (US) envoltades per inverted repeats. Aquestes repeticions permeten reordenaments de les regions úniques de forma que el tamany del genoma d'aquests virus pot variar fina a 10kpb.
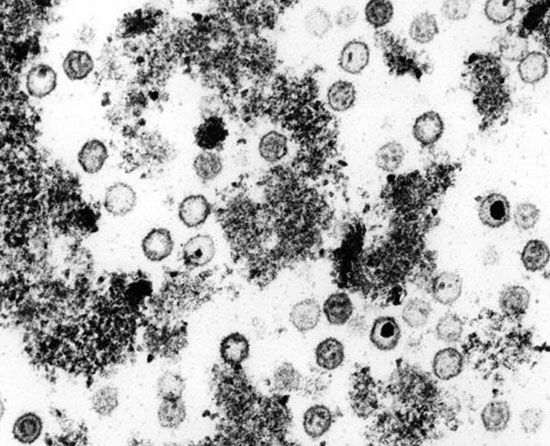
Per veure una fotografia de microscopia electrònica de HHV-6 clica aquí
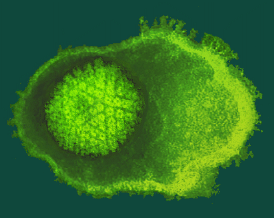
És el vuitè herpes virus humà i el més recentment identificat. Va ser aïllat de mostres de pacients amb VIH i sarcoma de Kaposi (KS) l'any 1994 per Chang et al. També es troba associat a limfoma primari de cavitats i en la malaltia de Castleman.
Fotografia de HHV-8:
1. Sarcoma de Kaposi:
És la neoplasia més freqüent en pacients amb SIDA. Es caracteritza per l'aparició de taques morades a nivell cutani de diferents tamanys, tot i que hi ha casos en que es presenta a nivell de ganglis i mucoses de la boca i intestí. Si el sarcoma afecta només a la pell no produeix la mort però si es dissemina a altes òrgans (pulmó, fetge, cor, ganglis linfàtics i cervell) pot produir la mort. La malaltia té diferents estadis:
En les lesions primerenques només un 10% de les cèl.lules endotelials i fusiformes són HHV-8 positives mentres que en els estadis posteriors al voltant del 90% estan infectades. Això suggereix que el virus confereix alguna avantatge en el creixement de les cèl.lules.
2. Limfoma primari de cavitats (PEL):
Es manifesta com efusions malignes en les cavitats de la pleura, pericardi o al peritoneu sense provocar cap massa tumoral.
La majoria dels pacients són HIV positius amb una immunosupressió avançada i en el 80% dels casos contenen EBV latent. El mecanisme molecular pel qual provoca la malaltia és desconegut.
La majoria de la recerca en KSHV es fa utilitzant línies cel.lulars en infecció latent establertes a partir de PEL.
3. Malaltia de Castleman (MCD):
Va ser descrita per primer cop als anys 50. És un síndrome limfoproliferatiu molt rar. Afecta a una varietat de teixits limfoides i no limfoides provocant febre, infiltració esplènica.
MCD és més freqüent en VIH positius i es presenta associat a Sarcoma de Kaposi.
Des del punt de vista clínic es manifesta com una massa local, normalment mediastínica encara que pot tenir altres localitzacions o ser generalitzada (multicèntrica).
4. Altres processos:
Existeixen controvèrsies respecte si el HHV-8 es troba present en teixit prostràtic i al tracte urogenital en individus sans. També s'ha descrit en cèl.lules dendrítiques de la medul.la en pacients amb mieloma. També s'ha identificat en lesions cutànies en pacients amb immunodeficiència no portadors de VIH i en alguns tumors vasculars.
Partim d'una seqüència de DNA d'herpesvirus associat a Sarcoma de Kaposi extreta de Genbank (entrada U75698, posicions 73021-85260) en format FASTA. Utilitzant el programa Translate predim totes les proteïnes codificades per la seqüència en els sis marcs de lectura. Hem escollit que la longitud mínima de les proteïnes que predim sigui de 60 aminoàcids.
Per a cada proteïna predita hem fet un protein-protein BLAST (blastp) per identificar les proteïnes de HHV-8 i trobar possibles proteïnes homòlogues en altres organismes.
Amb les seqüències proteiques que tenen una homologia més gran a la nostra hem decidit fer un alineament de seqüències amb el ClustalW per tal de poder veure quines són les regions més conservades i que, per tant, poden ser importants per a la funció de la proteïna. En l'apartat de resultats hem posat un exemple de l'alineament en el ClustalW.
1. Explicació del programa:
Introduïm una seqüència en format fasta i el nostre programa fa la complementària i la posa 5' 3'. Un cop tenim la seqüència forward i la reverse fem les tres pautes de lectura de cadescuna obtenint sis seqüències en sis variables diferents.
Per cada seqüència cridem dues funcions: la primera funció (atg) ens emmagatzema en un vector les possibles proteïnes, considerant l'inici el codó ATG i el final els codons STOP (TAA,TGA,TAG). A més, també emmagatzema en la variable $inici el nucleòtid d'inici i final de la proteïna. La segona funció, a partir del vector on tenim les seqüències proteiques, talla la seqüència per codons i mitjançant dos hashes obtenim les proteïnes. El primer hash (%codi) passa els codons a aminoàcid amb el codi de tres lletres i el segon hash (%lletra) passa el codi de tres lletres al d'una lletra, adjudicant al codó STOP un *.
Així obtenim una variable ($resultat) amb totes les possibles proteïnes separades per un asterisc. Amb la funció split tallem pels asteriscs i obtenim les proteïnes separades. Després les imprimim en un arxiu de sortida on tenim una línia d'informació de la proteïna i la seqüència en format fasta.
El programa permet triar l'allargada mínima que volem que tinguin les proteïnes que prediu.
Per veure com funciona més detalladament, consulteu el link del programa Translate on estan explicats tots els passos.
2. Abans d' utilitzar el programa hem de tenir en compte que:
>Prot1_pautalectura1_fwd_inici_final
Prot indica el número de proteïna
pautadelectura és un número del 1 al 3 que indica la pauta de lectura
fwd o rev segons si llegim la cadena forward o la reverse
inici_final número on comença i acaba la seqüència codificant
3. Com utilitzar el programa?
nomprograma.pl sequencia.fa arxiusortida.fa tall
sequencia.fa és la seqüència en format fasta que volem analitzar
arxiusortida.fa és l'arxiu on aniran a parar tots els resultats
tall és el número mínim d'aminoàcids que ha de tenir la proteïna
Per veure de forma clara els resultats obtinguts en el protein-protein BLAST els hem posat en una taula .
En aquesta taula veiem els resultats del BLAST de les diferents proteïnes que hem obtingut amb el nostre programa.
Condicions del blast: Blast contra tots els organismes, amb la matriu BLOSUM62, threshold=0.005.
Nomées incluim en la taula proteïnes amb un score superior a 50.
Resultats obtinguts amb el ClustalW per la proteïna dUTPase utilitzant les seqüències (amb l'entrada de Genbank) amb un score superior a 50.
Resultats obtinguts amb el Clustalw per la proteïna primase-helicase utilitzant les seqüències (amb l'entrada de Genbank) amb un score superior a 50.
El programa Translate ha predit un total de 32 proteïnes, 15 en forward i 17 en reverse. Utilitzant el BLAST hem pogut comfirmar l'existència de 10 d'aquestes:
Forward:
Reverse:
La majoria de proteïnes predites no han estat descrites anteriorment. El més probable és que les de menor longitud no existeixin ja que hem observat que les descrites solen ser de major tamany (són totes de longitud major de 100 aa). Per això, creiem que podrïem posar el mínim d'aa a 100.
Per assegurar-nos que les proteïnes predites realment es troben en les posicions predites les hem buscat al NCBI Entrez Gene. Posem un exemple de la proteïna EBV BLRF1 homolog a continuació. La nostre proteïna és la de color granate que està en reverse.
Respecte a la cerca de proteïnes homòlogues en altres organismes hem de destacar que la majoria les hem trobat en altres herpesvirus. Això és lògic ja que els virus tenen una elevada taxa de mutació de manera que és difícil trobar homologies. De fet, si haguéssim fet la cerca amb nucleòtids en lloc d'aminoàcids el més probable és que no haguéssim trobat homologia ni amb altres herpesvirus degut a que la taxa de canvi nucleotídica és major que la proteica (el codi genètic és degenerat).
Les proteïnes que tenen més homòlegs són les següents:
Si mirem les seves funcions veiem que totes les proteïnes es troben relacionades amb processos de replicació, transcripció i traducció. Això és lògic ja que aquestes proteïnes tenen una funció essencial per la vida del virus i per tant estan sotmeses a una pressió evolutiva més gran.
Què podem dir de l'anàlisi amb el Clustalw? D'aquestes cinc proteïnes mostrem l'anàlisi de les dues que es troben més conservades, ja que sabem que tenen dominis conservats:
Tot i que les seqüències són força diferents veiem que hi ha certes regions que es troben molt conservades, no havent-hi cap canvi entre les seqüències. Això fa pensar que aquests aminoàcids estan implicats en la funció de la proteïna, ja sigui formant part del centre actiu o bé interaccionant amb el substrat (per exemple). Les links dels alineaments amb clustalw es troben en l'apartat de resultats.
Com hem pogut veure el programa prediu una gran quantitat de proteïnes, la majoria de les quals no estan descrites. Podem concloure que, tot i que el programa funciona, calen altres criteris per assegurar una bona predicció de les proteïnes.
Romo, N., Torras, N. (2004) Programa Translate.
McGeoch, D.J., Dolan, A., Ralph, A.C. (2000). Toward a comprehensive phylogeny for mammalian and avian herpesviruses. J Virol. 74:10401-10406
Jenner, R.G., Boshoff, C. (2002). The molecular pathology of Kaposi's sarcoma-associated herpesvirus. Biochim Biophys Acta 1602:1-22.
Jenner, R.G., Albà, M.M., Boshoff, C., Kellam, P. (2001). Kaposi's sarcoma-associated herpesvirus latent and lytic gene expression as revealed by DNA arrays. J Virol. 75:891-902.
Holzerlandt, R., Orengo, C., Kellam, P., Albà, M.M. (2002). Identification of new herpesvirus gene homologues in the human genome. Genome Research 12:1739-1748.
BLAST (http://www.ncbi.nlm.nih.gov/blast)
Clustalw: http://www.ebi.ac.uk/clustalw/
Pubmed: http:///www.ncbi.nlm.nih.gov/PubMed/medline.html
Swissprot: http://us.expasy.org/sprot/
Informació herpes virus: http://www.virology.net/garryfavweb12.html
http://herpesvirus.tripod.com/gallery/gallery.htm
http://www.tulane.edu/~dmsander/WWW/335/Herpesviruses.html
http://www.google.com
Webmaestro: http://www.ldc.usb.ve/~vtheok/webmaestro/
DISCUSSIÓ

BIBLIOGRAFIA